

Antiproliferative factor-induced changes in phosphorylation and palmitoylation of cytoskeleton-associated protein-4 regulate its nuclear translocation and DNA binding
- PMID: 22536245
- PMCID: PMC3320026
- DOI: 10.1155/2012/150918
Antiproliferative factor-induced changes in phosphorylation and palmitoylation of cytoskeleton-associated protein-4 regulate its nuclear translocation and DNA binding
Abstract
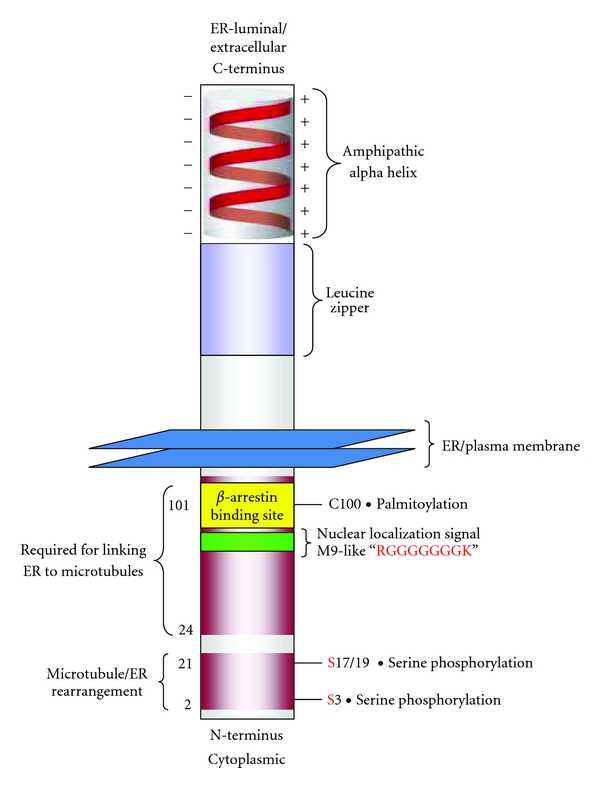
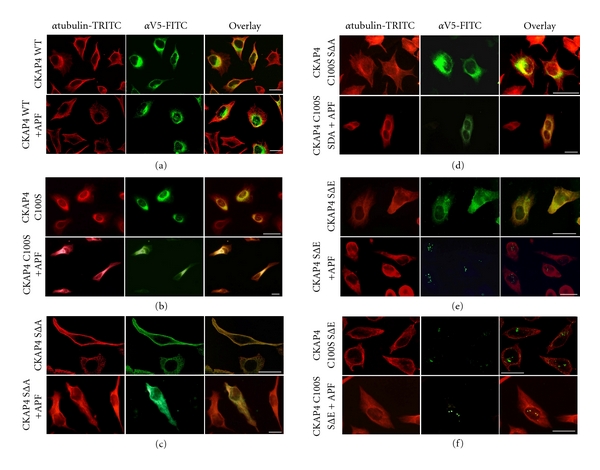
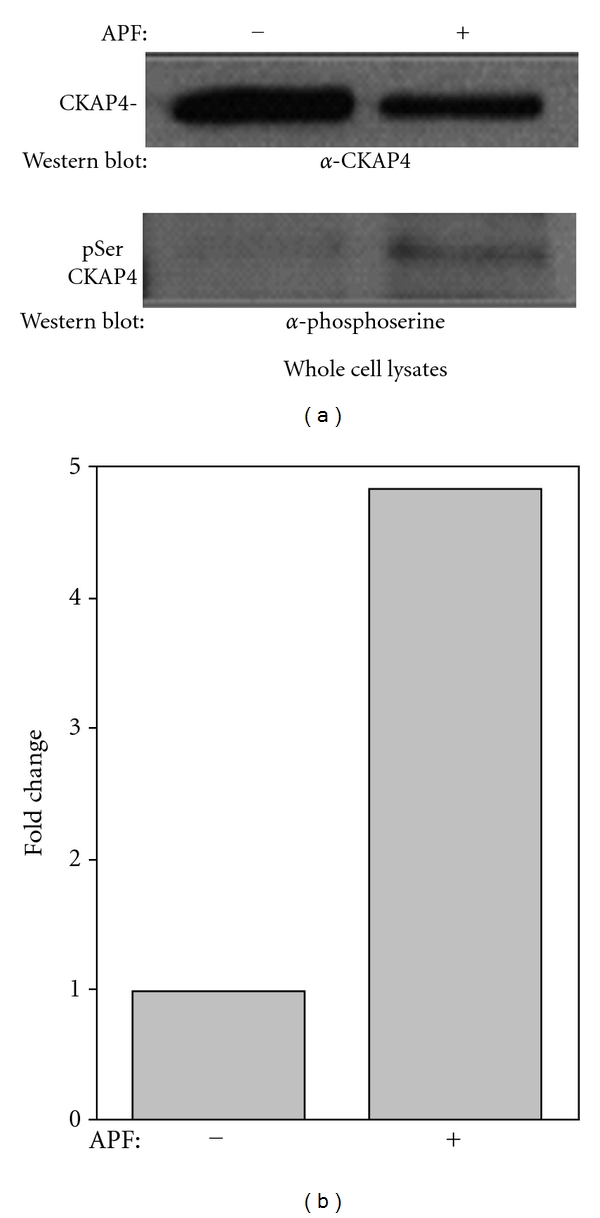
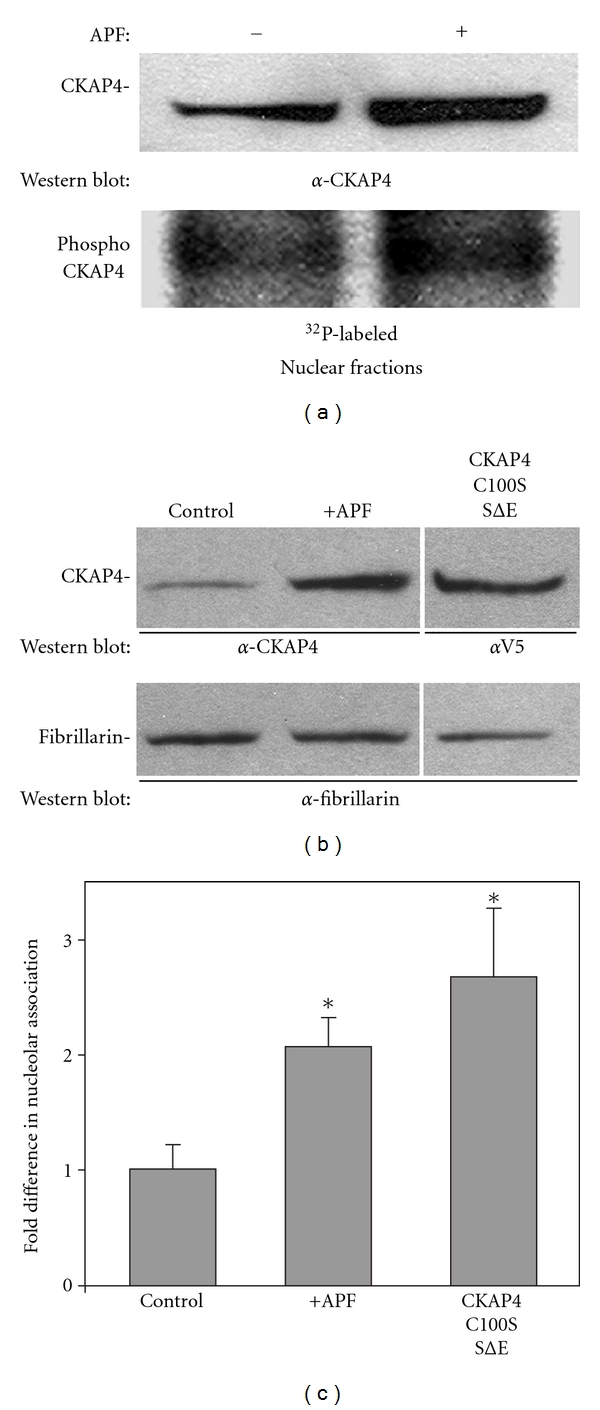
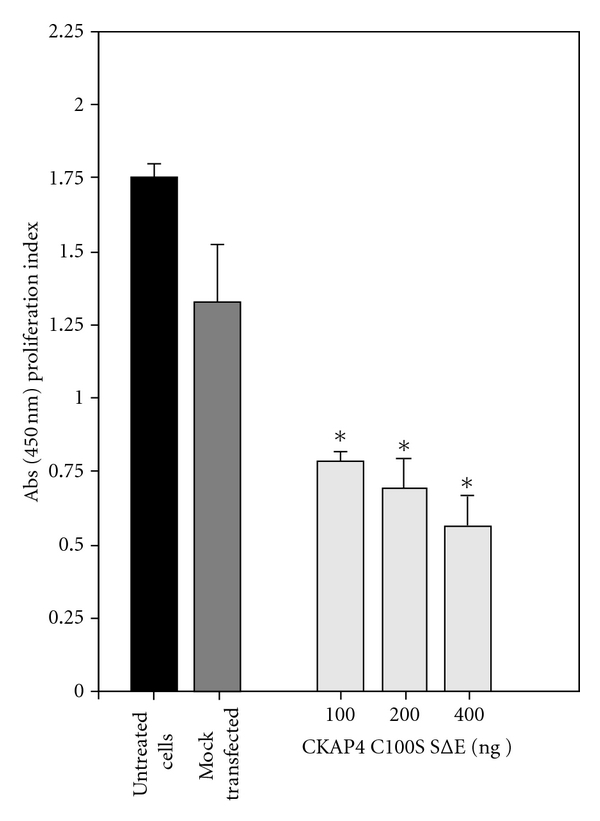
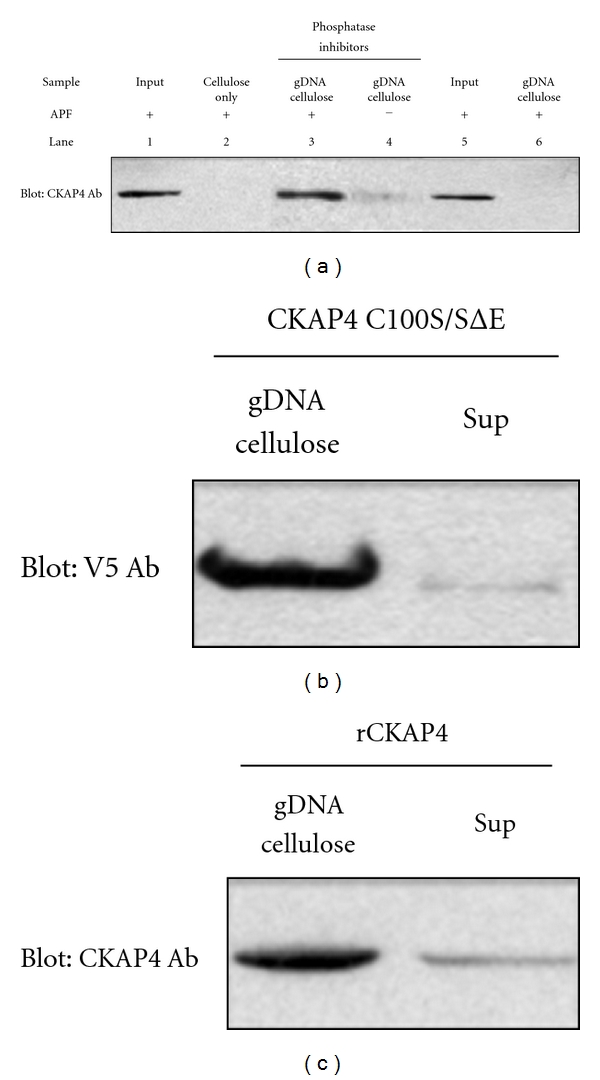
Cytoskeleton-associated protein 4 (CKAP4) is a reversibly palmitoylated and phosphorylated transmembrane protein that functions as a high-affinity receptor for antiproliferative factor (APF)-a sialoglycopeptide secreted from bladder epithelial cells of patients with interstitial cystitis (IC). Palmitoylation of CKAP4 by the palmitoyl acyltransferase, DHHC2, is required for its cell surface localization and subsequent APF signal transduction; however, the mechanism for APF signal transduction by CKAP4 is unknown. In this paper, we demonstrate that APF treatment induces serine phosphorylation of residues S3, S17, and S19 of CKAP4 and nuclear translocation of CKAP4. Additionally, we demonstrate that CKAP4 binds gDNA in a phosphorylation-dependent manner in response to APF treatment, and that a phosphomimicking, constitutively nonpalmitoylated form of CKAP4 localizes to the nucleus, binds DNA, and mimics the inhibitory effects of APF on cellular proliferation. These results reveal a novel role for CKAP4 as a downstream effecter for APF signal transduction.
Figures

References
-
- Schweizer A, Rohrer J, Jeno P, De Maio A, Buchman TG, Hauri HP. A reversibly palmitoylated resident protein (p63) of an ER-Golgi intermediate compartment is related to a circulatory shock resuscitation protein. Journal of Cell Science. 1993;104(3):685–694. - PubMed
-
- Schweizer A, Ericsson M, Bachi T, Griffiths G, Hauri HP. Characterization of a novel 63 kDa membrane protein. Implications for the organization of the ER-to-Golgi pathway. Journal of Cell Science. 1993;104(3):671–683. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
