

Kinetics of PKCε activating and inhibiting llama single chain antibodies and their effect on PKCε translocation in HeLa cells
- PMID: 22536418
- PMCID: PMC3334965
- DOI: 10.1371/journal.pone.0035630
Kinetics of PKCε activating and inhibiting llama single chain antibodies and their effect on PKCε translocation in HeLa cells
Abstract
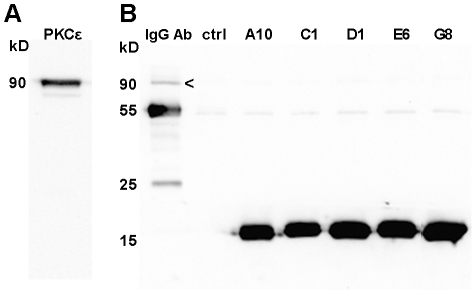
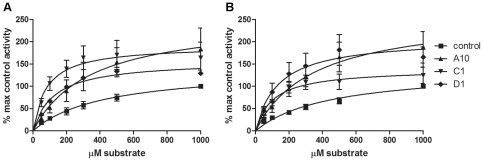
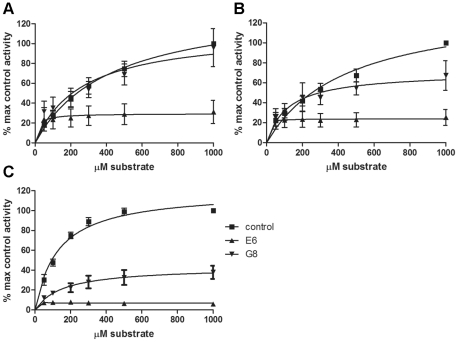
Dysregulation of PKCε is involved in several serious diseases such as cancer, type II diabetes and Alzheimer's disease. Therefore, specific activators and inhibitors of PKCε hold promise as future therapeutics, in addition to being useful in research into PKCε regulated pathways. We have previously described llama single chain antibodies (VHHs) that specifically activate (A10, C1 and D1) or inhibit (E6 and G8) human recombinant PKCε. Here we report a thorough kinetic analysis of these VHHs. The inhibiting VHHs act as non-competitive inhibitors of PKCε activity, whereas the activating VHHs have several different modes of action, either increasing V(max) and/or decreasing K(m) values. We also show that the binding of the VHHs to PKCε is conformation-dependent, rendering the determination of affinities difficult. Apparent affinities are in the micromolar range based on surface plasmon resonance studies. Furthermore, the VHHs have no effect on the activity of rat PKCε nor can they bind the rat form of the protein in immunoprecipitation studies despite the 98% identity between the human and rat PKCε proteins. Finally, we show for the first time that the VHHs can influence PKCε function also in cells, since an activating VHH increases the rate of PKCε translocation in response to PMA in HeLa cells, whereas an inhibiting VHH slows down the translocation. These results give insight into the mechanisms of PKCε activity modulation and highlight the importance of protein conformation on VHH binding.
Conflict of interest statement
Figures




References
-
- Newton AC. Regulation of protein kinase C. Curr Opin Cell Biol. 1997;9(2):161–167. - PubMed
-
- Liu WS, Heckman CA. The sevenfold way of PKC regulation. Cell Signal. 1998;10(8):529–542. - PubMed
-
- Akita Y. Protein kinase C-epsilon (PKC-epsilon): Its unique structure and function. J Biochem. 2002;132(6):847–852. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
