

The Nkx5/HMX homeodomain protein MLS-2 is required for proper tube cell shape in the C. elegans excretory system
- PMID: 22537498
- PMCID: PMC3358442
- DOI: 10.1016/j.ydbio.2012.03.015
The Nkx5/HMX homeodomain protein MLS-2 is required for proper tube cell shape in the C. elegans excretory system
Abstract
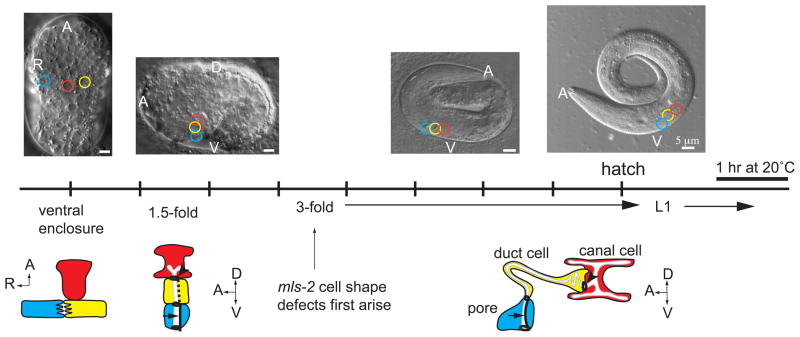
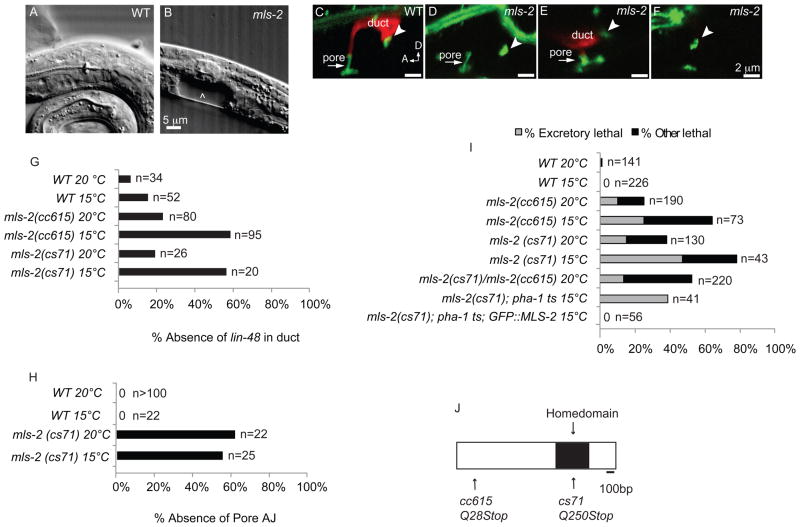
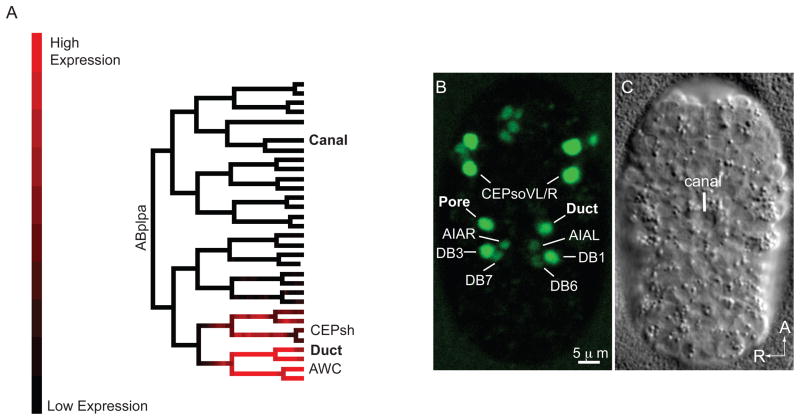
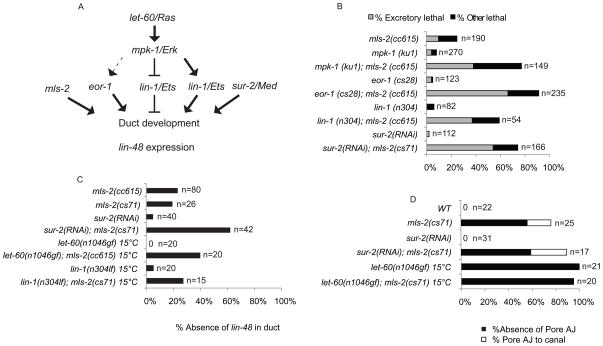
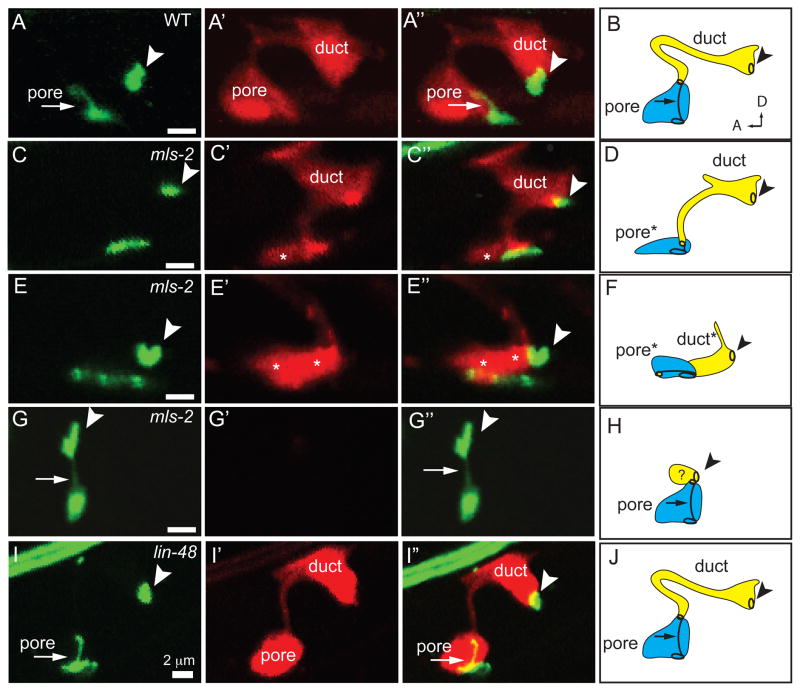
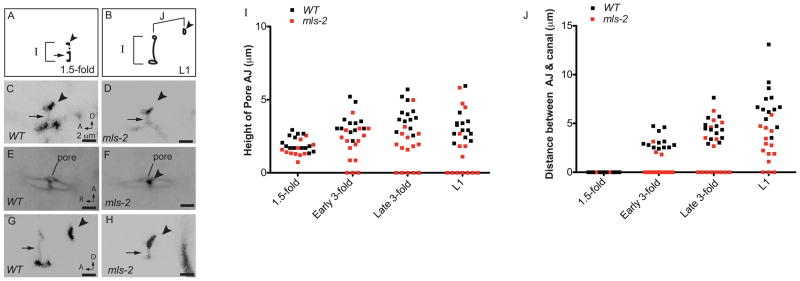
Cells perform wide varieties of functions that are facilitated, in part, by adopting unique shapes. Many of the genes and pathways that promote cell fate specification have been elucidated. However, relatively few transcription factors have been identified that promote shape acquisition after fate specification. Here we show that the Nkx5/HMX homeodomain protein MLS-2 is required for cellular elongation and shape maintenance of two tubular epithelial cells in the C. elegans excretory system, the duct and pore cells. The Nkx5/HMX family is highly conserved from sea urchins to humans, with known roles in neuronal and glial development. MLS-2 is expressed in the duct and pore, and defects in mls-2 mutants first arise when the duct and pore normally adopt unique shapes. MLS-2 cooperates with the EGF-Ras-ERK pathway to turn on the LIN-48/Ovo transcription factor in the duct cell during morphogenesis. These results reveal a novel interaction between the Nkx5/HMX family and the EGF-Ras pathway and implicate a transcription factor, MLS-2, as a regulator of cell shape.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
The Caenorhabditis elegans Excretory System: A Model for Tubulogenesis, Cell Fate Specification, and Plasticity.Genetics. 2016 May;203(1):35-63. doi: 10.1534/genetics.116.189357. Genetics. 2016. PMID: 27183565 Free PMC article. Review.
-
The HMX/NKX homeodomain protein MLS-2 specifies the identity of the AWC sensory neuron type via regulation of the ceh-36 Otx gene in C. elegans.Development. 2010 Mar;137(6):963-74. doi: 10.1242/dev.044719. Epub 2010 Feb 11. Development. 2010. PMID: 20150279 Free PMC article.
-
The HMX homeodomain protein MLS-2 regulates cleavage orientation, cell proliferation and cell fate specification in the C. elegans postembryonic mesoderm.Development. 2005 Sep;132(18):4119-30. doi: 10.1242/dev.01967. Epub 2005 Aug 17. Development. 2005. PMID: 16107479
-
Synergistic roles of homeodomain proteins UNC-62 homothorax and MLS-2 HMX/NKX in the specification of olfactory neurons in Caenorhabditis elegans.Genetics. 2021 Oct 2;219(2):iyab133. doi: 10.1093/genetics/iyab133. Genetics. 2021. PMID: 34849889 Free PMC article.
-
Signal transduction and cell fate specification during Caenorhabditis elegans vulval development.Curr Opin Genet Dev. 1994 Aug;4(4):508-16. doi: 10.1016/0959-437x(94)90065-b. Curr Opin Genet Dev. 1994. PMID: 7950317 Review.
Cited by
-
The Bicoid class homeodomain factors ceh-36/OTX and unc-30/PITX cooperate in C. elegans embryonic progenitor cells to regulate robust development.PLoS Genet. 2015 Mar 4;11(3):e1005003. doi: 10.1371/journal.pgen.1005003. eCollection 2015 Mar. PLoS Genet. 2015. PMID: 25738873 Free PMC article.
-
A lineage-resolved molecular atlas of C. elegans embryogenesis at single-cell resolution.Science. 2019 Sep 20;365(6459):eaax1971. doi: 10.1126/science.aax1971. Epub 2019 Sep 5. Science. 2019. PMID: 31488706 Free PMC article.
-
A dimerized HMX1 inhibits EPHA6/epha4b in mouse and zebrafish retinas.PLoS One. 2014 Jun 19;9(6):e100096. doi: 10.1371/journal.pone.0100096. eCollection 2014. PLoS One. 2014. PMID: 24945320 Free PMC article.
-
mNG-tagged mls-2 knock-in alleles in C. elegans.MicroPubl Biol. 2022 Feb 22;2022:10.17912/micropub.biology.000529. doi: 10.17912/micropub.biology.000529. eCollection 2022. MicroPubl Biol. 2022. PMID: 35224463 Free PMC article.
-
The Caenorhabditis elegans Excretory System: A Model for Tubulogenesis, Cell Fate Specification, and Plasticity.Genetics. 2016 May;203(1):35-63. doi: 10.1534/genetics.116.189357. Genetics. 2016. PMID: 27183565 Free PMC article. Review.
References
-
- Bargmann CI, Hartwieg E, Horvitz HR. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 1993;74:515–27. - PubMed
-
- Beitel GJ, Tuck S, Greenwald I, Horvitz HR. The Caenorhabditis elegans gene lin-1 encodes an ETS-domain protein and defines a branch of the vulval induction pathway. Genes Dev. 1995;9:3149–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
