

Zinc maintains prophase I arrest in mouse oocytes through regulation of the MOS-MAPK pathway
- PMID: 22539682
- PMCID: PMC3406555
- DOI: 10.1095/biolreprod.112.099390
Zinc maintains prophase I arrest in mouse oocytes through regulation of the MOS-MAPK pathway
Abstract
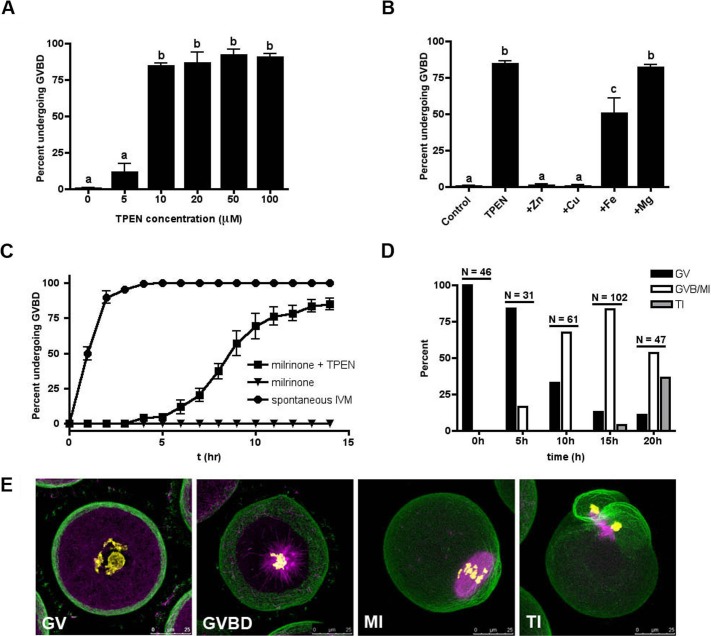
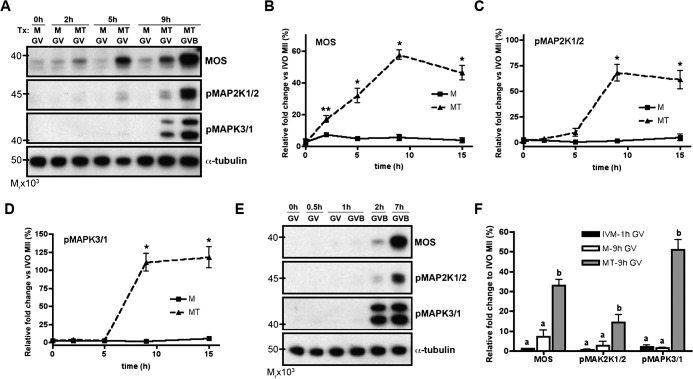
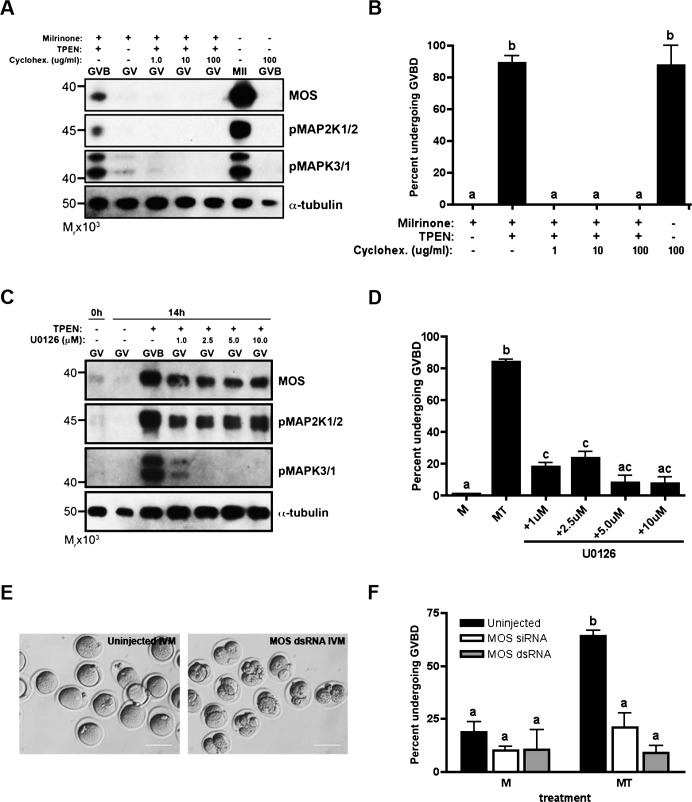
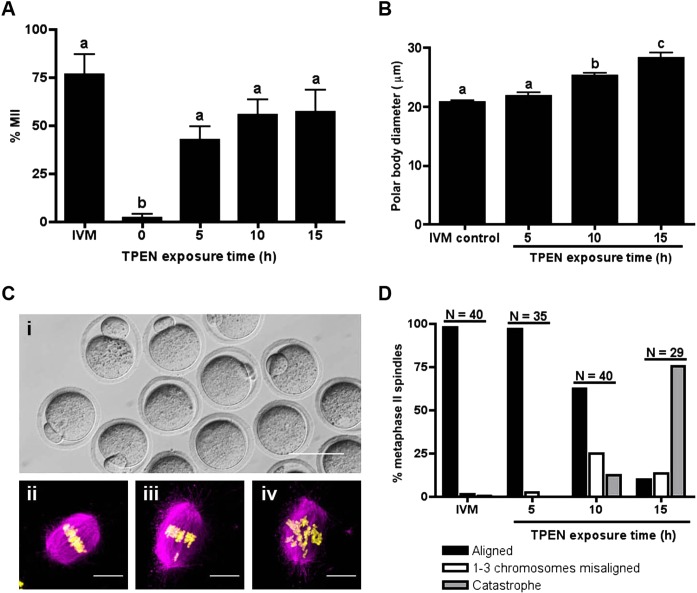
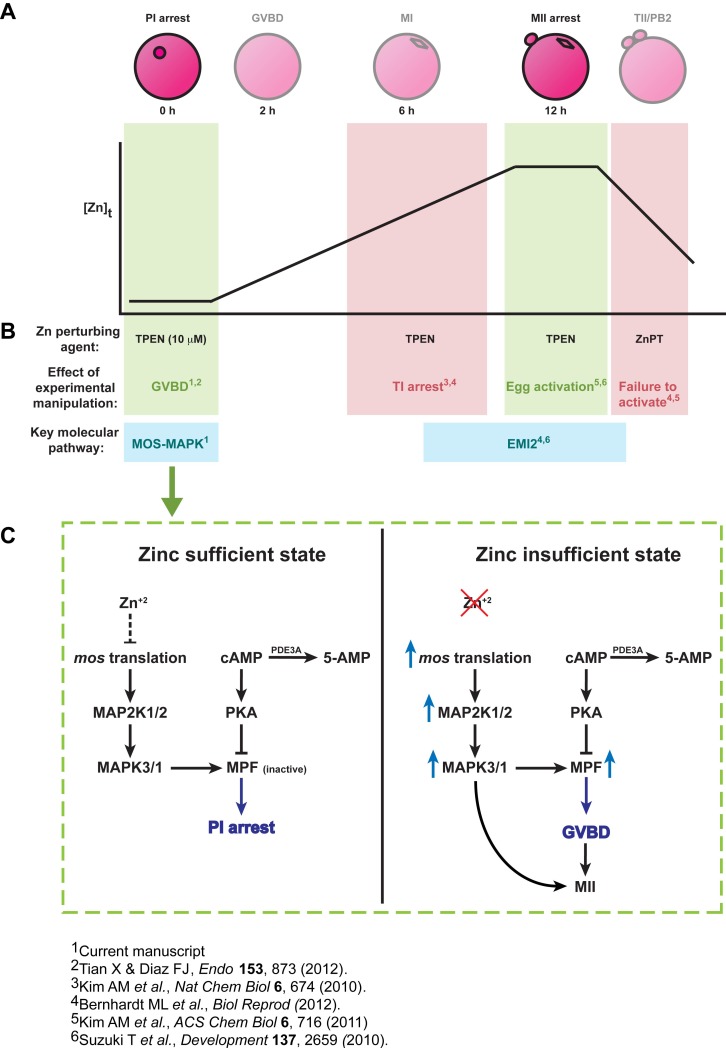
Meiosis in mammalian females is marked by two arrest points, at prophase I and metaphase II, which must be tightly regulated in order to produce a haploid gamete at the time of fertilization. The transition metal zinc has emerged as a necessary and dynamic regulator of the establishment, maintenance, and exit from metaphase II arrest, but the roles of zinc during prophase I arrest are largely unknown. In this study, we investigate the mechanisms of zinc regulation during the first meiotic arrest. Disrupting zinc availability in the prophase I arrested oocyte by treatment with the heavy metal chelator N,N,N',N'-tetrakis-(2-pyridylmethyl)-ethylenediamine (TPEN) causes meiotic resumption even in the presence of pharmacological inhibitors of meiosis. We further show that the MOS-MAPK pathway mediates zinc-dependent prophase I arrest, as the pathway prematurely activates during TPEN-induced meiotic resumption. Conversely, inhibition of the MOS-MAPK pathway maintains prophase I arrest. While prolonged zinc insufficiency ultimately results in telophase I arrest, early and transient exposure of oocytes to TPEN is sufficient to induce meiotic resumption and bypass the telophase I block, allowing the formation of developmentally competent eggs upon parthenogenetic activation. These results establish zinc as a crucial regulator of meiosis throughout the entirety of oocyte maturation, including the maintenance of and release from the first and second meiotic arrest points.
Figures
References
-
- Tsafriri A. Mammalian oocyte maturation: model systems and their physiological relevance. Adv Exp Med Biol 1979; 112: 269 281 - PubMed
-
- Josefsberg LB, Galiani D, Lazar S, Kaufman O, Seger R, Dekel N. Maturation-promoting factor governs mitogen-activated protein kinase activation and interphase suppression during meiosis of rat oocytes. Biol Reprod 2003; 68: 1282 1290 - PubMed
-
- Conti M, Andersen CB, Richard F, Mehats C, Chun SY, Horner K, Jin C, Tsafriri A. Role of cyclic nucleotide signaling in oocyte maturation. Mol Cell Endocrinol 2002; 187: 153 159 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
