

The subthalamic nucleus is one of multiple innervation sites for long-range corticofugal axons: a single-axon tracing study in the rat
- PMID: 22539859
- PMCID: PMC3479642
- DOI: 10.1523/JNEUROSCI.5717-11.2012
The subthalamic nucleus is one of multiple innervation sites for long-range corticofugal axons: a single-axon tracing study in the rat
Abstract
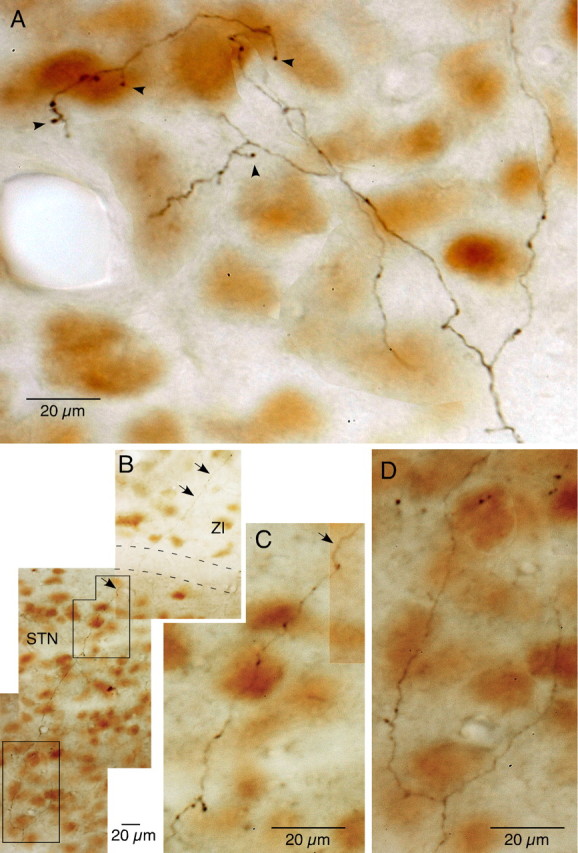
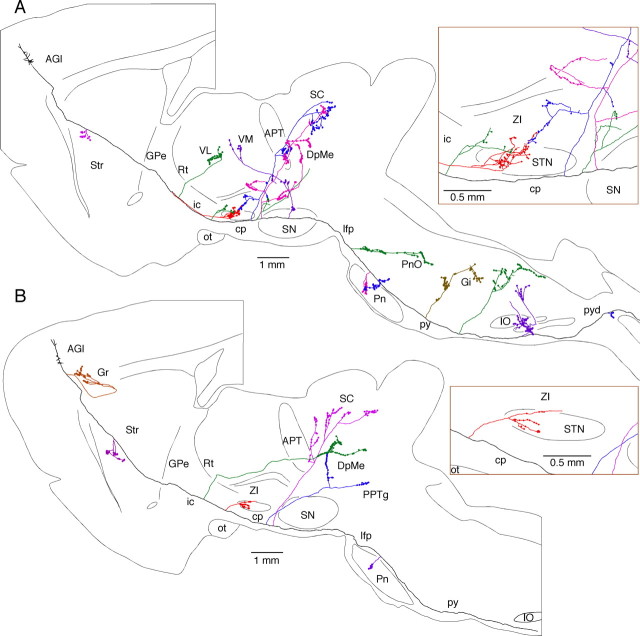
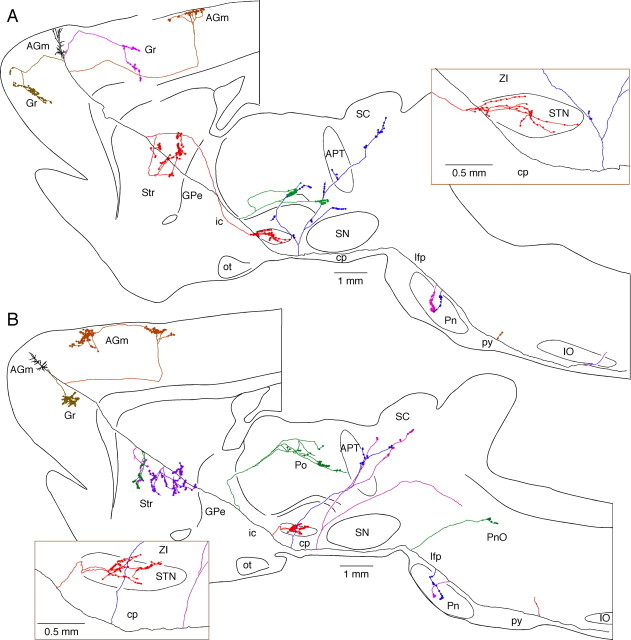
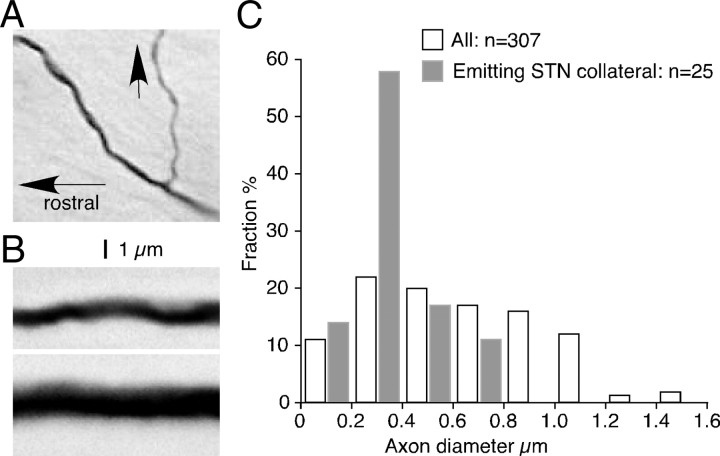
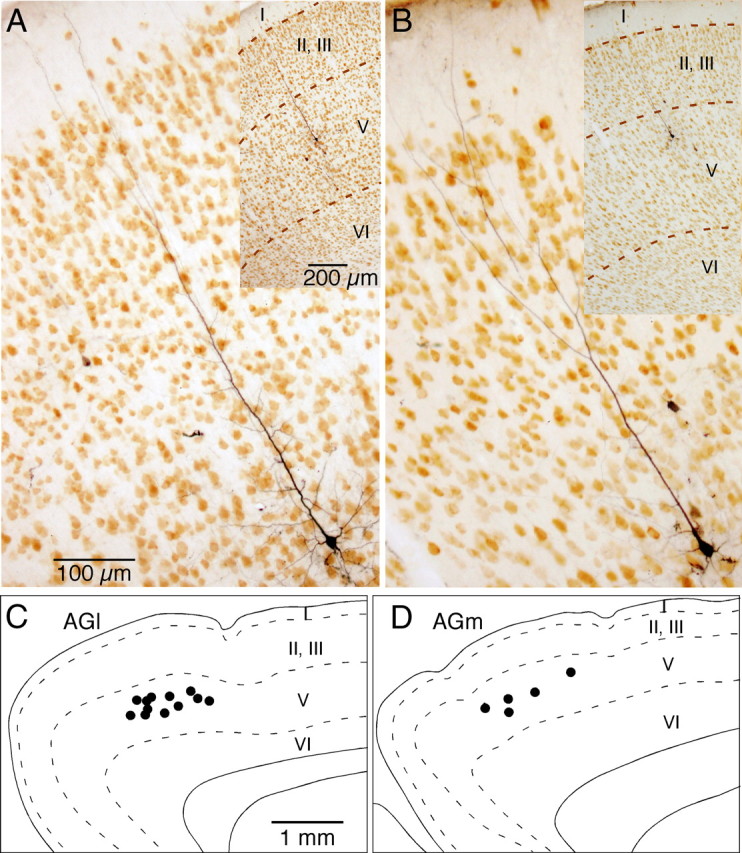
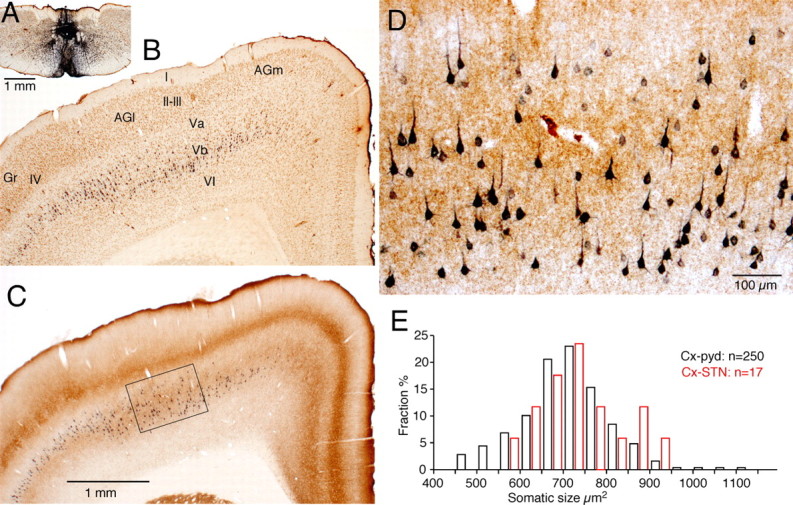
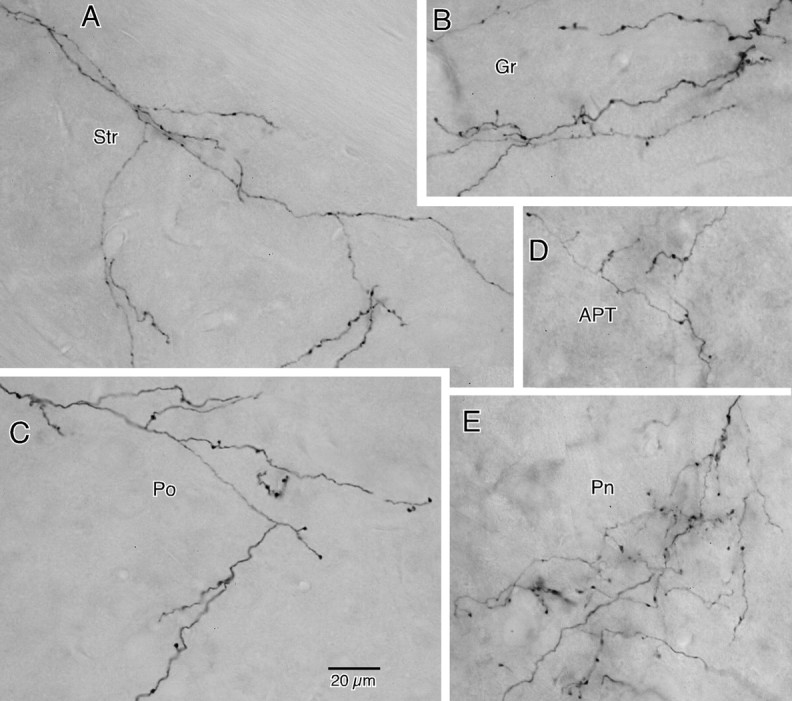
The frontal cortex provides strong excitatory inputs to the subthalamic nucleus (STN), and these cortico-STN inputs play critical roles in the control of basal ganglia activity. It has been assumed from anatomical and physiological studies that STN is innervated mainly by collaterals of thick and fast conducting pyramidal tract axons originating from the frontal cortex deep layer V neurons, implying that STN directly receives efferent copies of motor commands. To more closely examine this assumption, we performed biotinylated dextran amine anterograde tracing studies in rats to examine the cortical layer of origin, the sizes of parent axons, and whether or not the cortical axons emit any other collaterals to brain areas other than STN. This study revealed that the cortico-STN projection is formed mostly by collaterals of a small fraction of small-to-medium-sized long-range corticofugal axons, which also emit collaterals that innervate multiple other brain sites including the striatum, associative thalamic nuclei, superior colliculus, zona incerta, pontine nucleus, multiple other brainstem areas, and the spinal cord. The results imply that some layer V neurons are involved in associative control of movement through multiple brain innervation sites and that the cortico-STN projection is one part of this multiple corticofugal system.
Figures
References
-
- Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci. 1989;12:366–375. - PubMed
-
- Bevan MD, Francis CM, Bolam JP. The glutamate-enriched cortical and thalamic input to neurons in the subthalamic nucleus of the rat: convergence with GABA-positive terminals. J Comp Neurol. 1995;361:491–511. - PubMed
-
- Brecht M, Krauss A, Muhammad S, Sinai-Esfahani L, Bellanca S, Margrie TW. Organization of rat vibrissa motor cortex and adjacent areas according to cytoarchitectonics, microstimulation, and intracellular stimulation of identified cells. J Comp Neurol. 2004;479:360–373. - PubMed
-
- Canteras NS, Shammah-Lagnado SJ, Silva BA, Ricardo JA. Afferent connections of the subthalamic nucleus: a combined retrograde and anterograde horseradish peroxidase study in the rat. Brain Res. 1990;513:43–59. - PubMed
-
- Chang HT, Kuo H, Whittaker JA, Cooper NG. Light and electron microscopic analysis of projection neurons retrogradely labeled with Fluoro-Gold: notes on the application of antibodies to Fluoro-Gold. J Neurosci Methods. 1990;35:31–37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources