

Nitric oxide modulates bacterial biofilm formation through a multicomponent cyclic-di-GMP signaling network
- PMID: 22542454
- PMCID: PMC3361614
- DOI: 10.1016/j.molcel.2012.03.023
Nitric oxide modulates bacterial biofilm formation through a multicomponent cyclic-di-GMP signaling network
Abstract
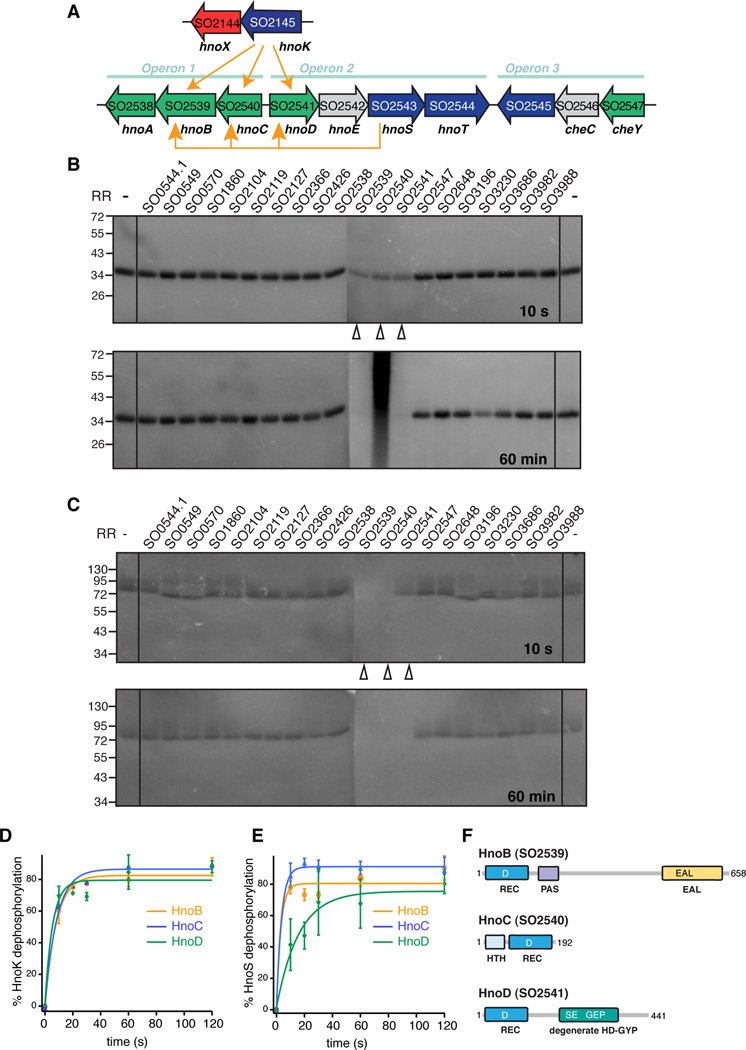
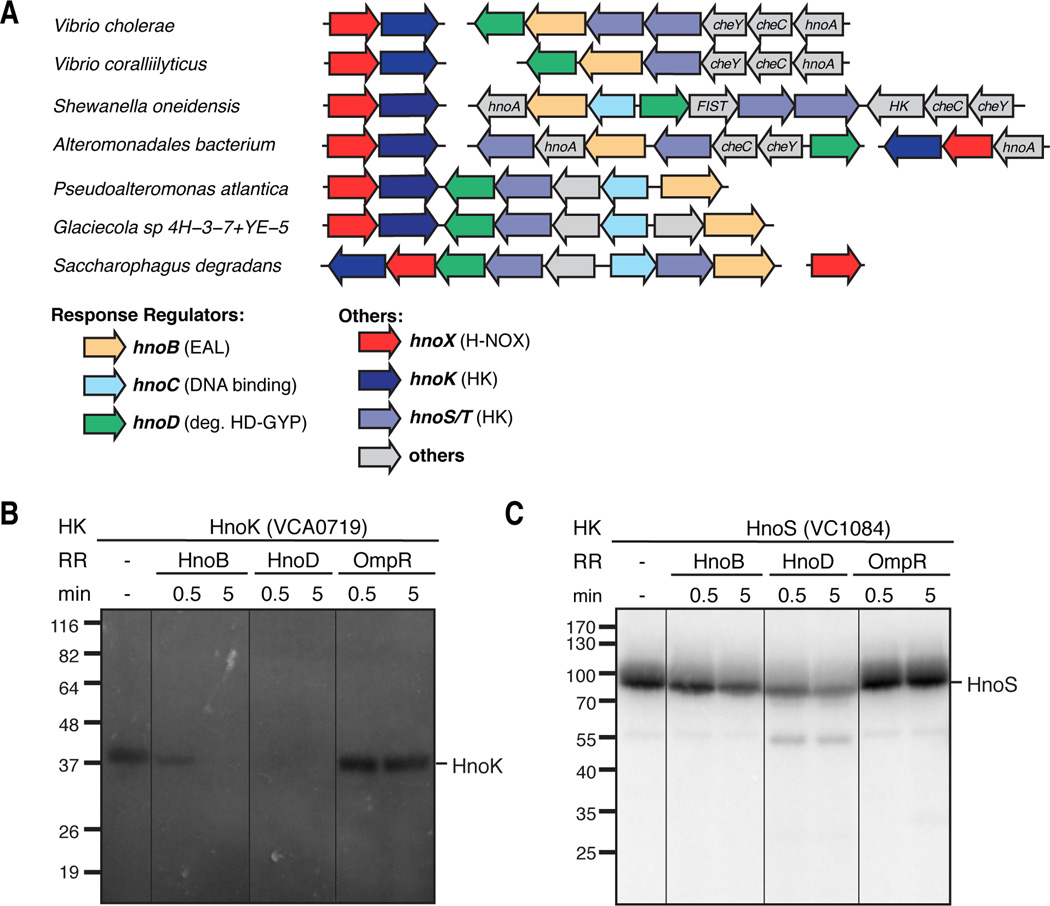
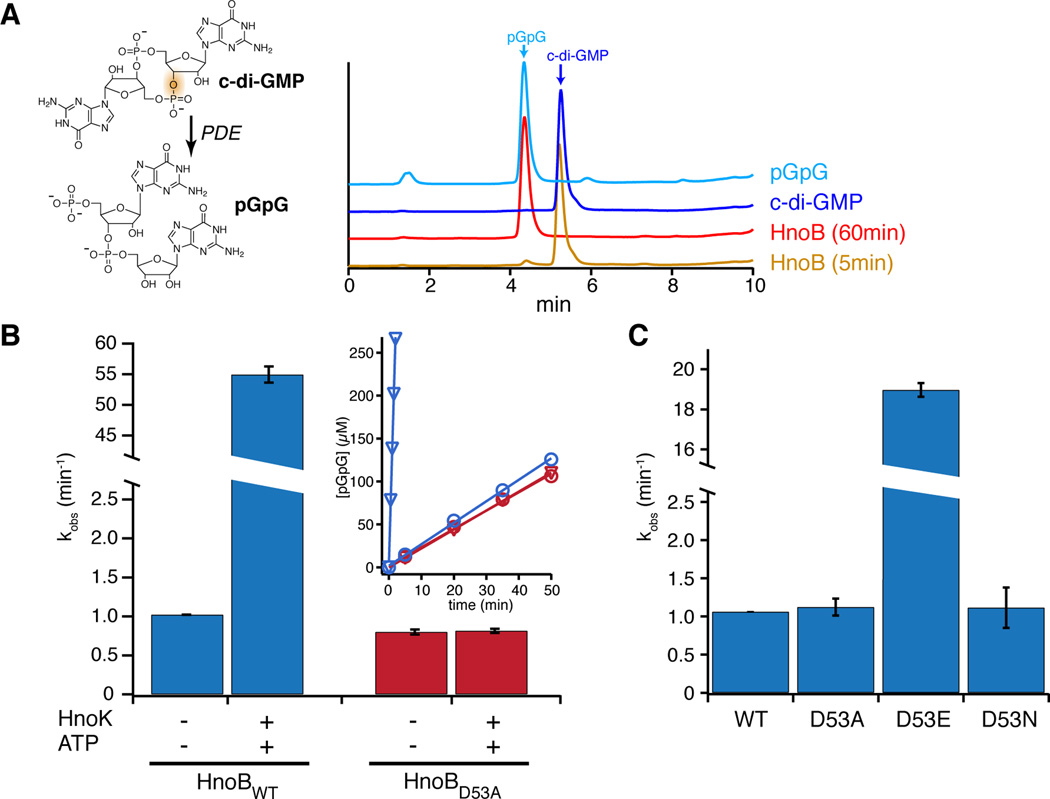
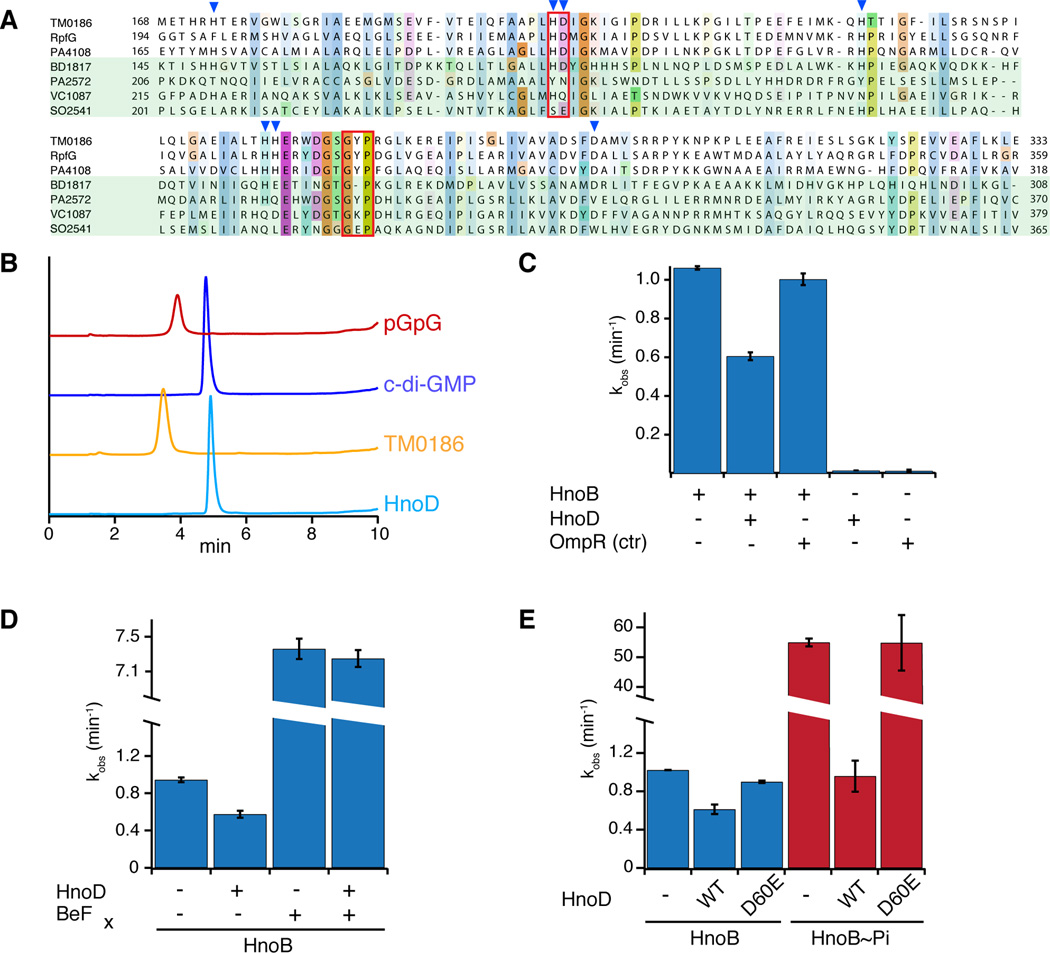
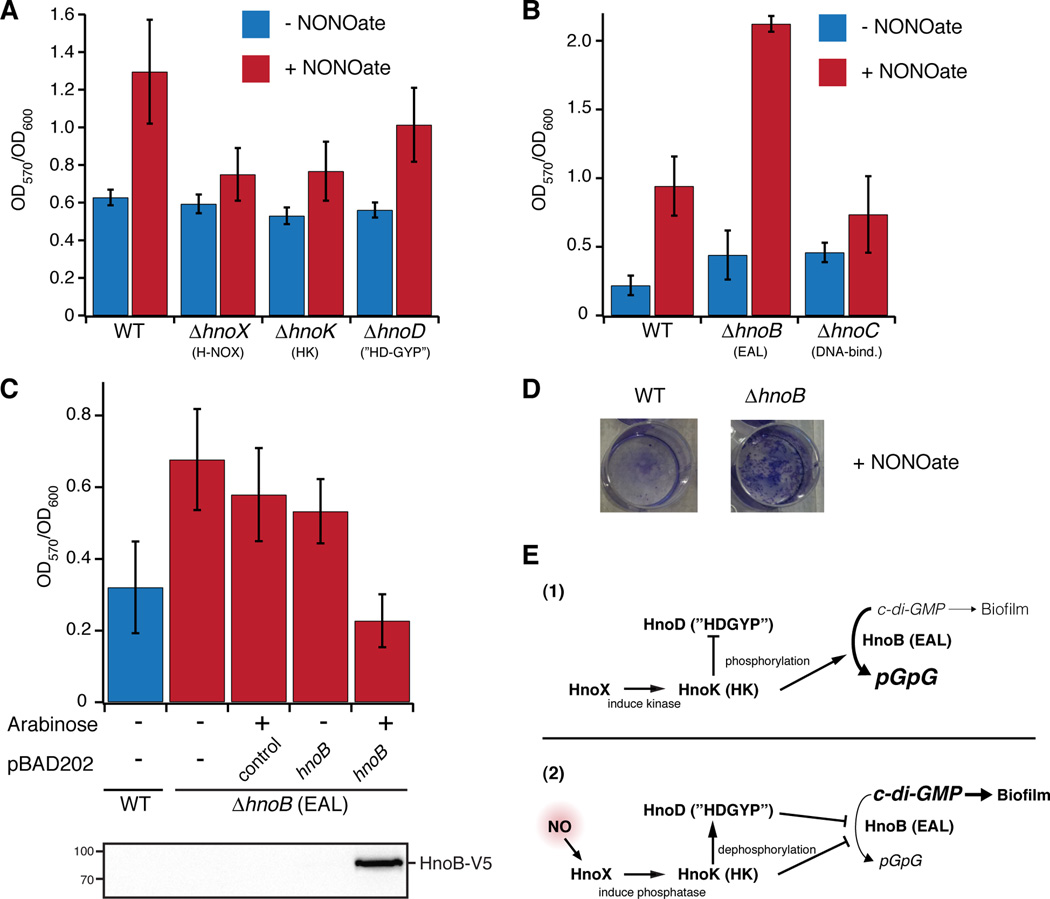
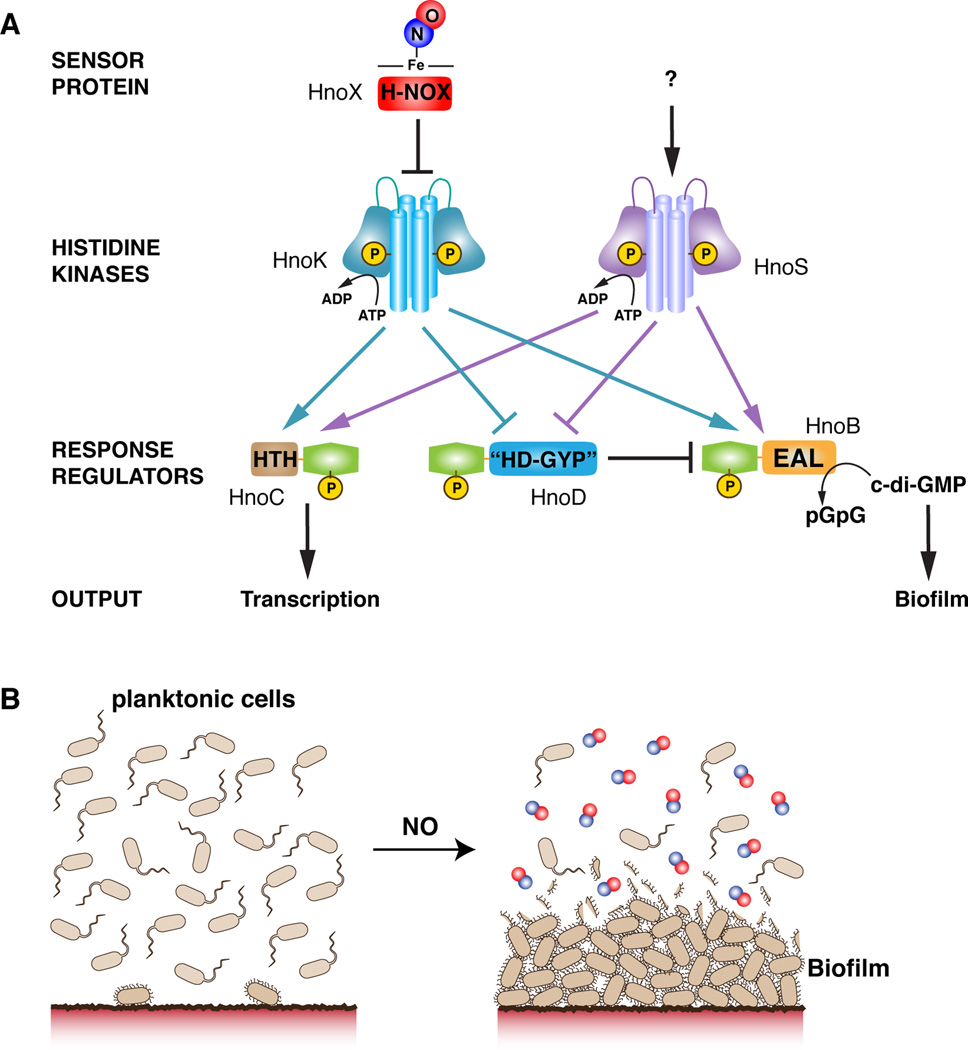
Nitric oxide (NO) signaling in vertebrates is well characterized and involves the heme-nitric oxide/oxygen-binding (H-NOX) domain of soluble guanylate cyclase as a selective NO sensor. In contrast, little is known about the biological role or signaling output of bacterial H-NOX proteins. Here, we describe a molecular pathway for H-NOX signaling in Shewanella oneidensis. NO stimulates biofilm formation by controlling the levels of the bacterial secondary messenger cyclic diguanosine monophosphate (c-di-GMP). Phosphotransfer profiling was used to map the connectivity of a multicomponent signaling network that involves integration from two histidine kinases and branching to three response regulators. A feed-forward loop between response regulators with phosphodiesterase domains and phosphorylation-mediated activation intricately regulated c-di-GMP levels. Phenotypic characterization established a link between NO signaling and biofilm formation. Cellular adhesion may provide a protection mechanism for bacteria against reactive and damaging NO. These results are broadly applicable to H-NOX-mediated NO signaling in bacteria.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Barends TRM, Hartmann E, Griese JJ, Beitlich T, Kirienko NV, Ryjenkov DA, Reinstein J, Shoeman RL, Gomelsky M, Schlichting I. Structure and mechanism of a bacterial light-regulated cyclic nucleotide phosphodiesterase. Nature. 2009;459:1015–1018. - PubMed
-
- Boehm A, Kaiser M, Li H, Spangler C, Kasper CA, Ackermann M, Kaever V, Sourjik V, Roth V, Jenal U. Second messenger-mediated adjustment of bacterial swimming velocity. Cell. 2010;141:107–116. - PubMed
-
- Boon EM, Davis JH, Tran R, Karow DS, Huang SH, Pan D, Miazgowicz MM, Mathies RA, Marletta MA. Nitric oxide binding to prokaryotic homologs of the soluble guanylate cyclase beta1 H-NOX domain. J. Biol. Chem. 2006;281:21892–21902. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
