

Morphogenetic fields in embryogenesis, regeneration, and cancer: non-local control of complex patterning
- PMID: 22542702
- PMCID: PMC3413735
- DOI: 10.1016/j.biosystems.2012.04.005
Morphogenetic fields in embryogenesis, regeneration, and cancer: non-local control of complex patterning
Abstract
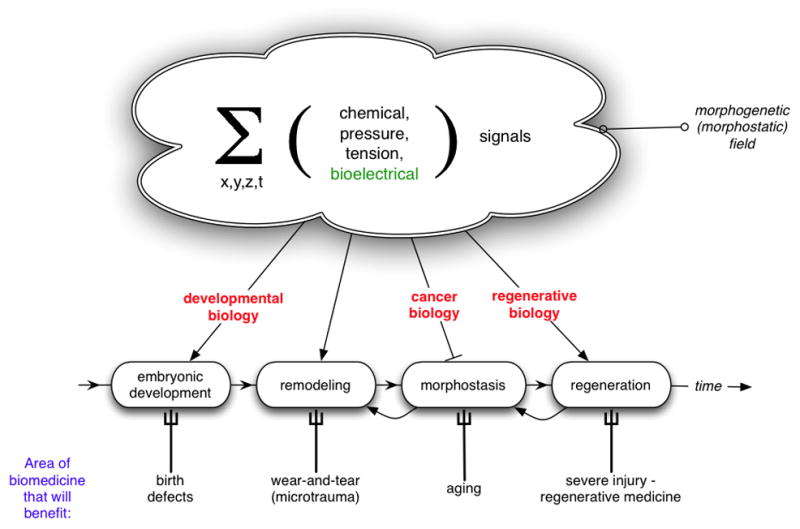
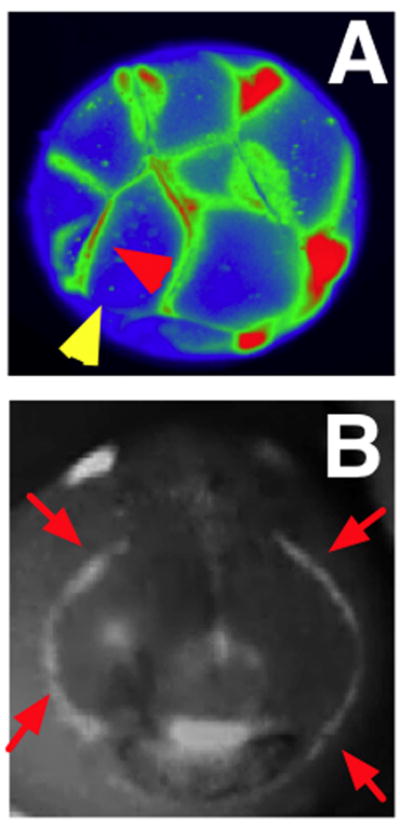
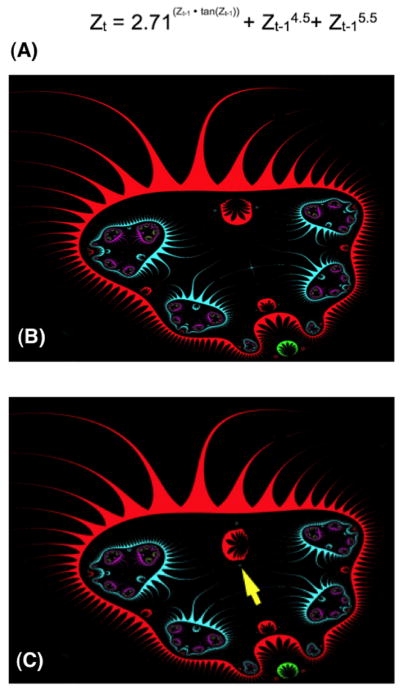
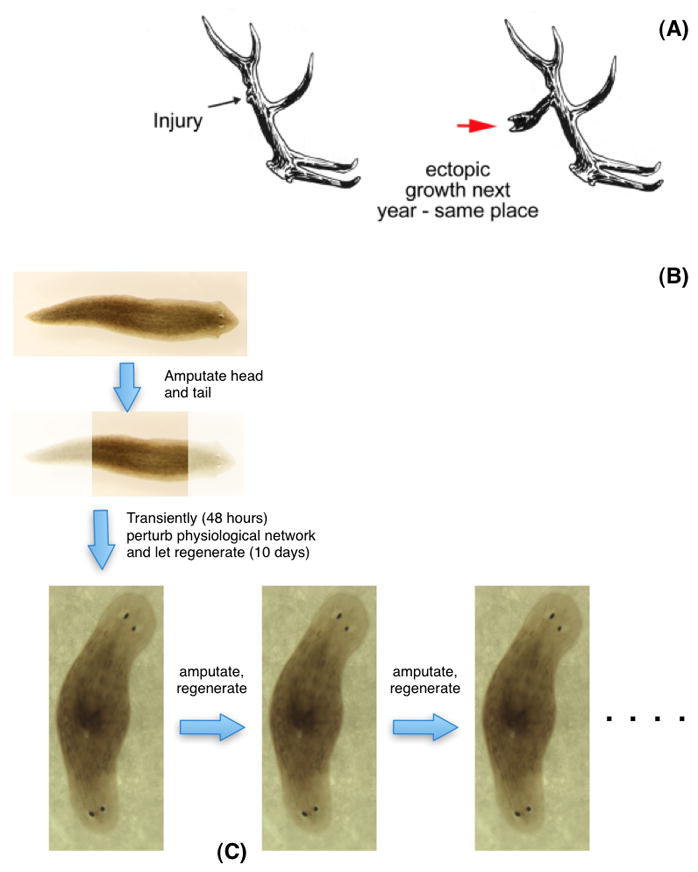
Establishment of shape during embryonic development, and the maintenance of shape against injury or tumorigenesis, requires constant coordination of cell behaviors toward the patterning needs of the host organism. Molecular cell biology and genetics have made great strides in understanding the mechanisms that regulate cell function. However, generalized rational control of shape is still largely beyond our current capabilities. Significant instructive signals function at long range to provide positional information and other cues to regulate organism-wide systems properties like anatomical polarity and size control. Is complex morphogenesis best understood as the emergent property of local cell interactions, or as the outcome of a computational process that is guided by a physically encoded map or template of the final goal state? Here I review recent data and molecular mechanisms relevant to morphogenetic fields: large-scale systems of physical properties that have been proposed to store patterning information during embryogenesis, regenerative repair, and cancer suppression that ultimately controls anatomy. Placing special emphasis on the role of endogenous bioelectric signals as an important component of the morphogenetic field, I speculate on novel approaches for the computational modeling and control of these fields with applications to synthetic biology, regenerative medicine, and evolutionary developmental biology.
Copyright © 2012 Elsevier Ireland Ltd. All rights reserved.
Figures
References
-
- Adamatzky A. Game of Life cellular automata. Springer; London: 2010. p. xix.p. 579.
-
- Adams DS, Masi A, Levin M. H+ pump-dependent changes in membrane voltage are an early mechanism necessary and sufficient to induce Xenopus tail regeneration. Development. 2007;134:1323–1335. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
