

Chromatin modulation at the FLO11 promoter of Saccharomyces cerevisiae by HDAC and Swi/Snf complexes
- PMID: 22542969
- PMCID: PMC3389975
- DOI: 10.1534/genetics.112.140301
Chromatin modulation at the FLO11 promoter of Saccharomyces cerevisiae by HDAC and Swi/Snf complexes
Abstract
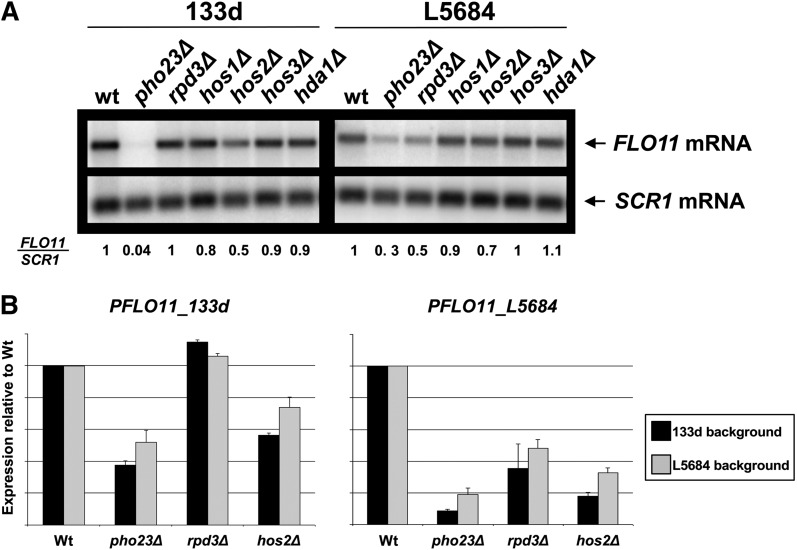
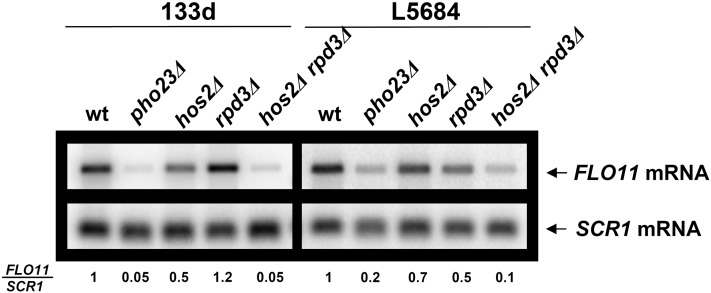
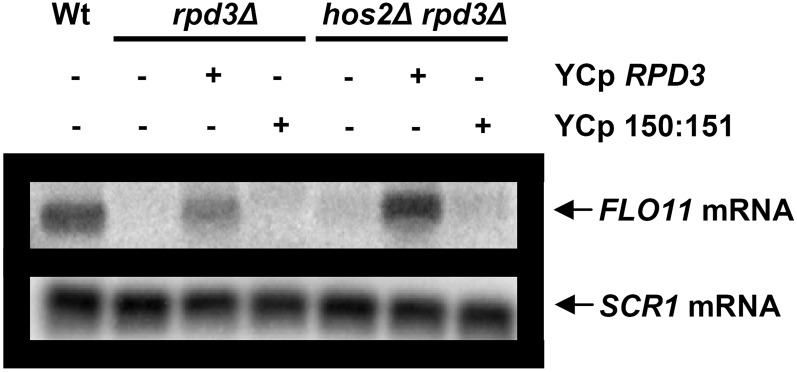
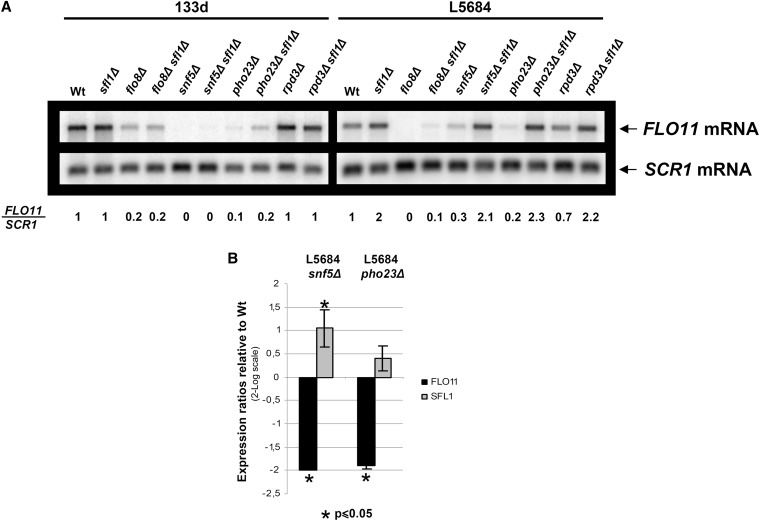
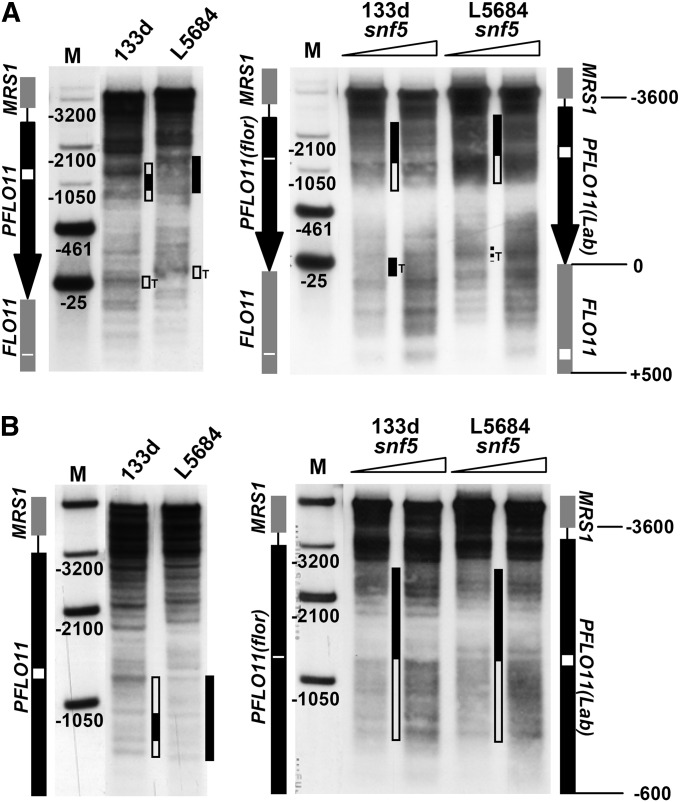
Cell adhesion and biofilm formation are critical processes in the pathogenicity of fungi and are mediated through a family of adhesin proteins conserved throughout yeasts and fungi. In Saccharomyces cerevisiae, Flo11 is the main adhesin involved in cell adhesion and biofilm formation, making the study of its function and regulation in this nonpathogenic budding yeast highly relevant. The S. cerevisiae FLO11 gene is driven by a TATA-box-containing promoter that is regulated through one of the longest regulatory upstream regions (3 kb) in yeast. We reported recently that two chromatin cofactor complexes, the Rpd3L deacetylase and the Swi/Snf chromatin-remodeling complexes, contribute significantly to the regulation of FLO11. Here, we analyze directly how these complexes impact on FLO11 promoter chromatin structure and dissect further the interplay between histone deacetylases, chromatin remodeling, and the transcriptional repressor Sfl1. We show that the regulation of chromatin structure represents an important layer of control in the highly complex regulation of the FLO11 promoter.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
