

TGD1, -2, and -3 proteins involved in lipid trafficking form ATP-binding cassette (ABC) transporter with multiple substrate-binding proteins
- PMID: 22544736
- PMCID: PMC3375562
- DOI: 10.1074/jbc.M112.370213
TGD1, -2, and -3 proteins involved in lipid trafficking form ATP-binding cassette (ABC) transporter with multiple substrate-binding proteins
Abstract
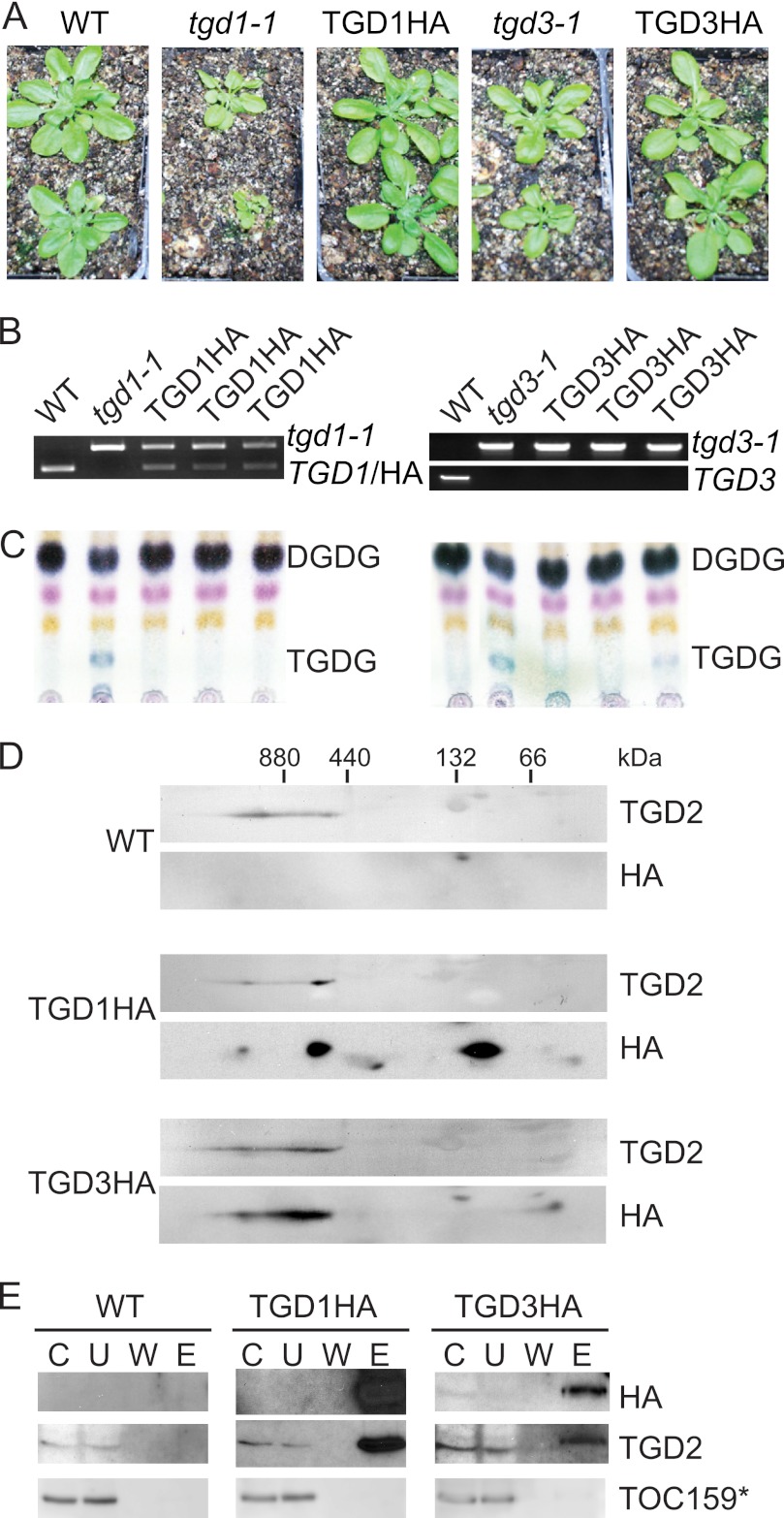
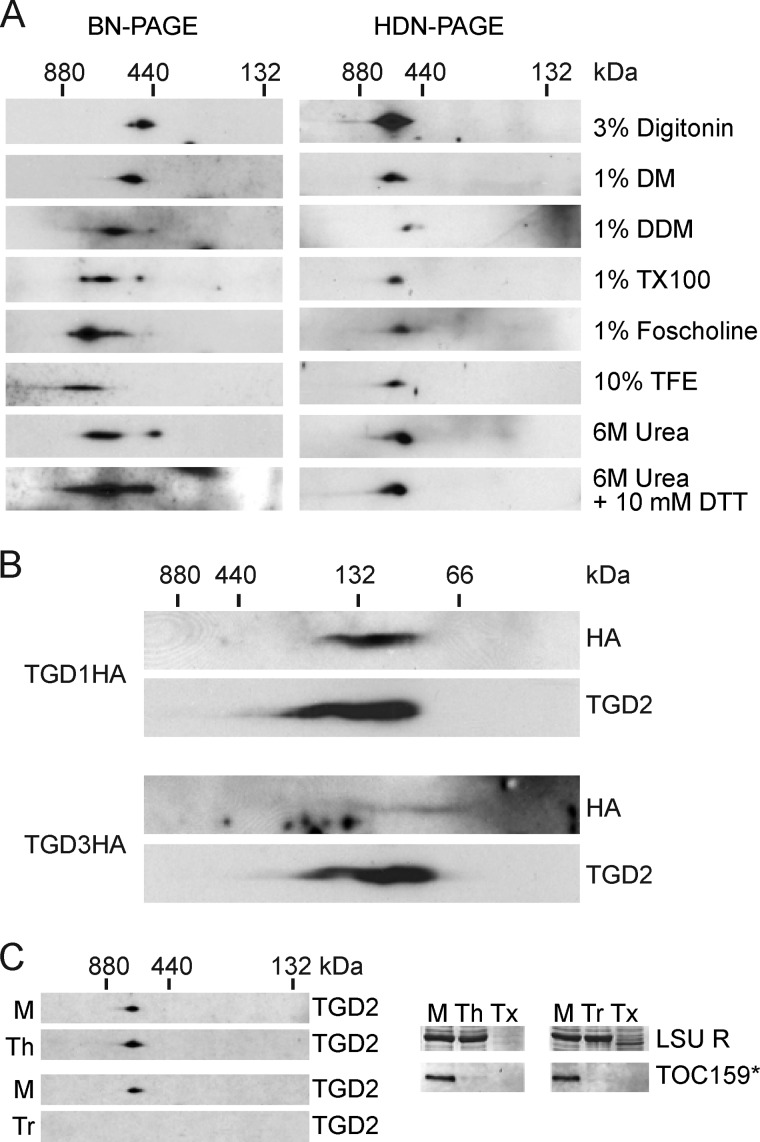
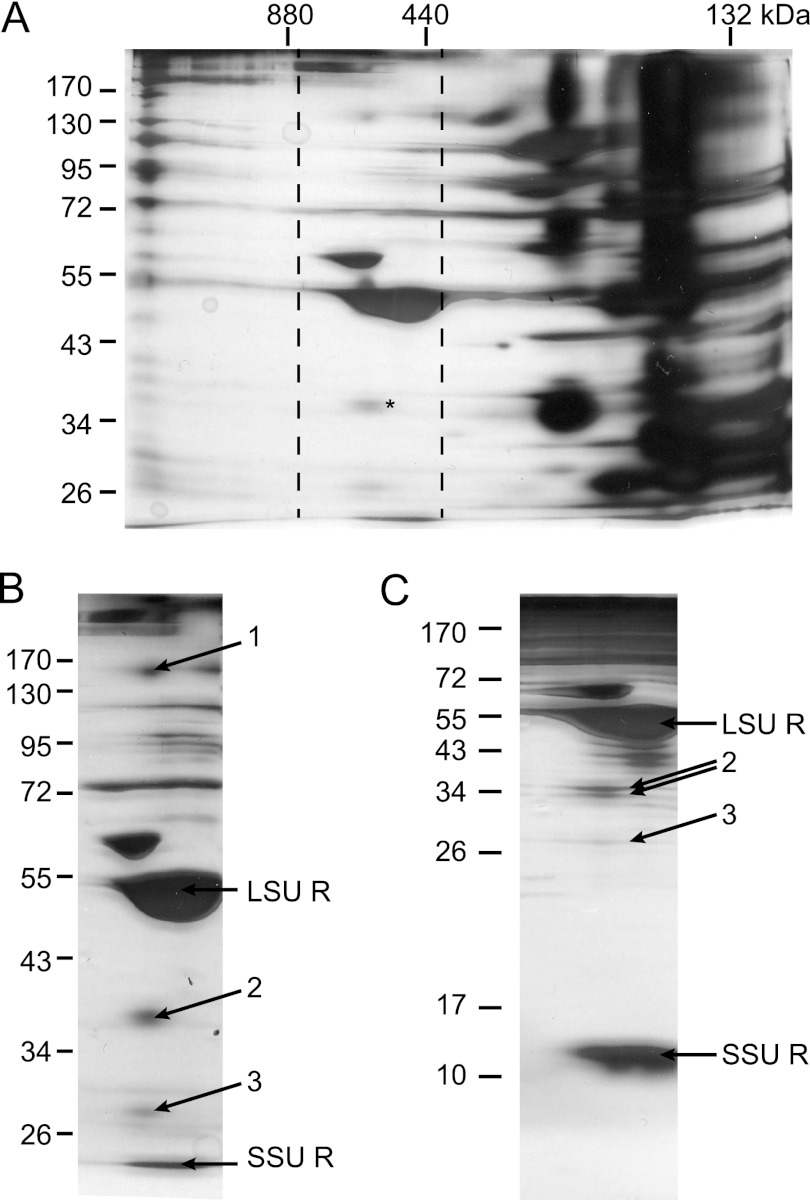
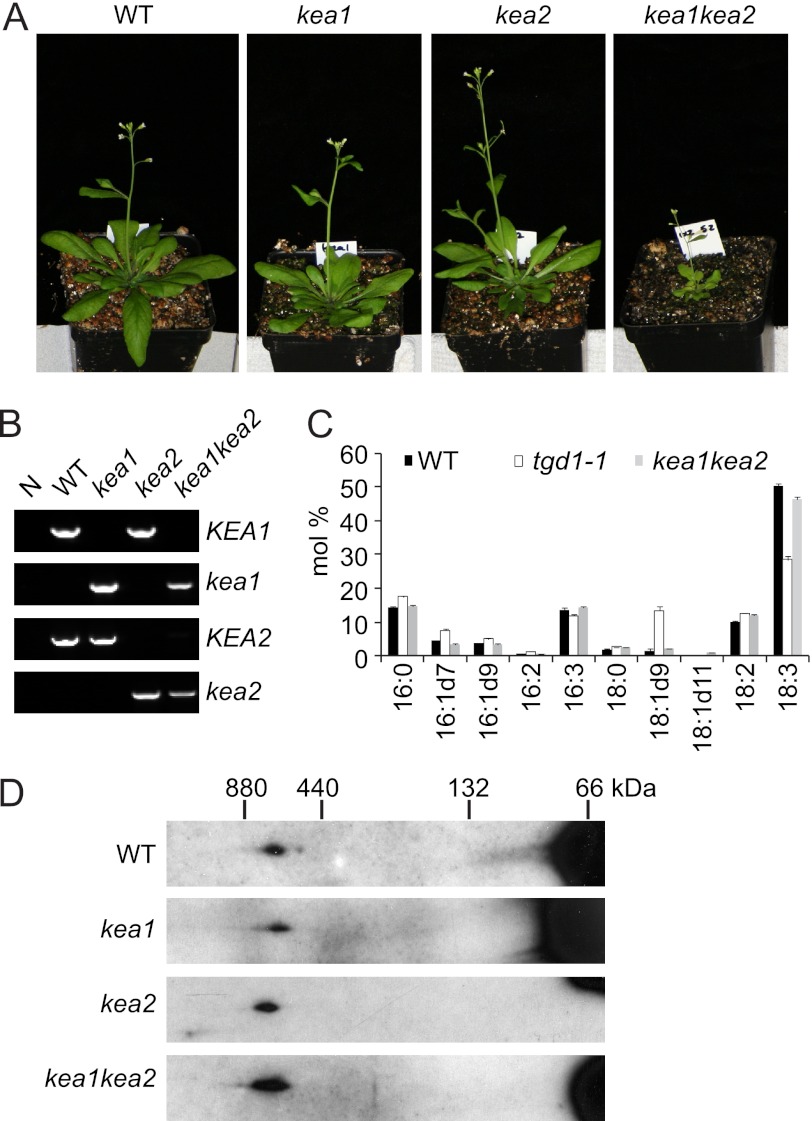
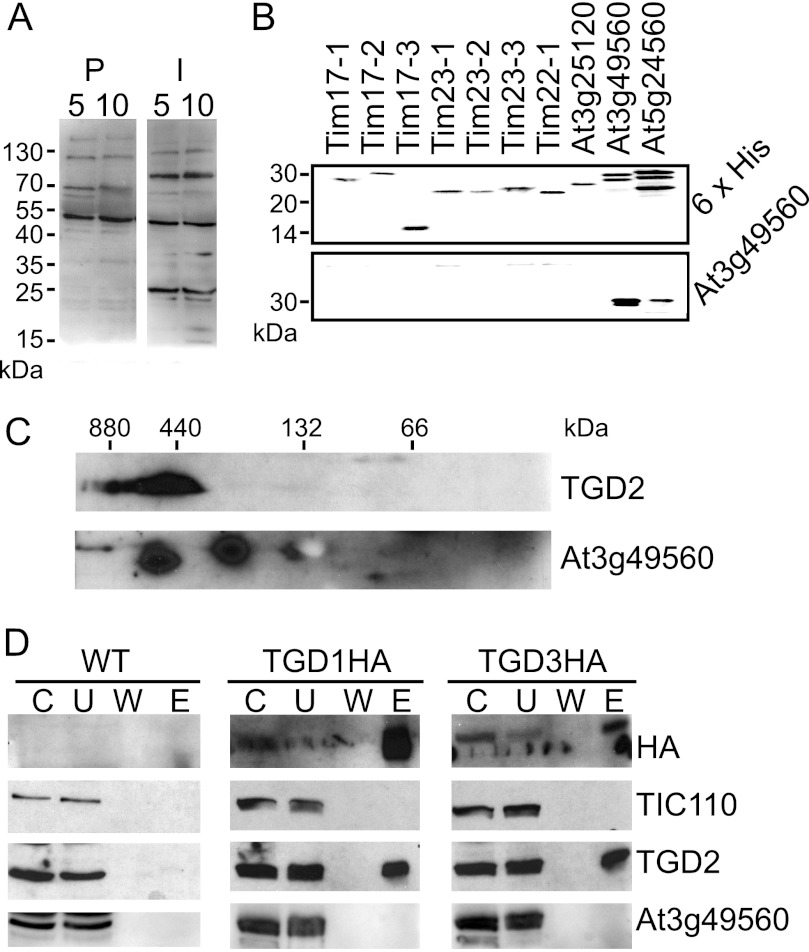
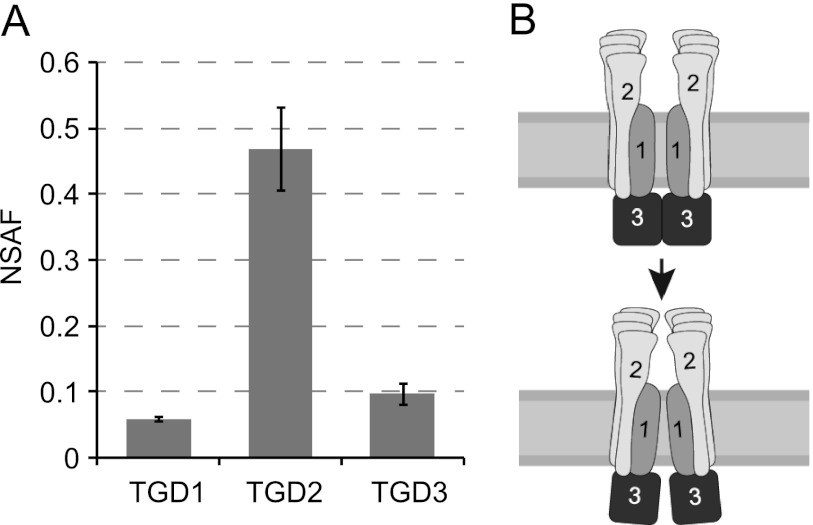
Members of the ATP-binding cassette (ABC) transporter family are essential proteins in species as diverse as archaea and humans. Their domain architecture has remained relatively fixed across these species, with rare exceptions. Here, we show one exception to be the trigalactosyldiacylglycerol 1, 2, and 3 (TGD1, -2, and -3) putative lipid transporter located at the chloroplast inner envelope membrane. TGD2 was previously shown to be in a complex of >500 kDa. We demonstrate that this complex also contains TGD1 and -3 and is very stable because it cannot be broken down by gentle denaturants to form a "core" complex similar in size to standard ABC transporters. The complex was purified from Pisum sativum (pea) chloroplast envelopes by native gel electrophoresis and examined by mass spectrometry. Identified proteins besides TGD1, -2, or -3 included a potassium efflux antiporter and a TIM17/22/23 family protein, but these were shown to be in separate high molecular mass complexes. Quantification of the complex components explained the size of the complex because 8-12 copies of the substrate-binding protein (TGD2) were found per functional transporter.
Figures
References
-
- Albers S. V., Koning S. M., Konings W. N., Driessen A. J. (2004) Insights into ABC transport in archaea. J. Bioenerg. Biomembr. 36, 5–15 - PubMed
-
- Higgins C. F. (1992) ABC transporters. From microorganisms to man. Annu. Rev. Cell Biol. 8, 67–113 - PubMed
-
- Hollenstein K., Dawson R. J., Locher K. P. (2007) Structure and mechanism of ABC transporter proteins. Curr. Opin. Struct. Biol. 17, 412–418 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
