

Function of Cryptococcus neoformans KAR7 (SEC66) in karyogamy during unisexual and opposite-sex mating
- PMID: 22544906
- PMCID: PMC3370462
- DOI: 10.1128/EC.00066-12
Function of Cryptococcus neoformans KAR7 (SEC66) in karyogamy during unisexual and opposite-sex mating
Abstract
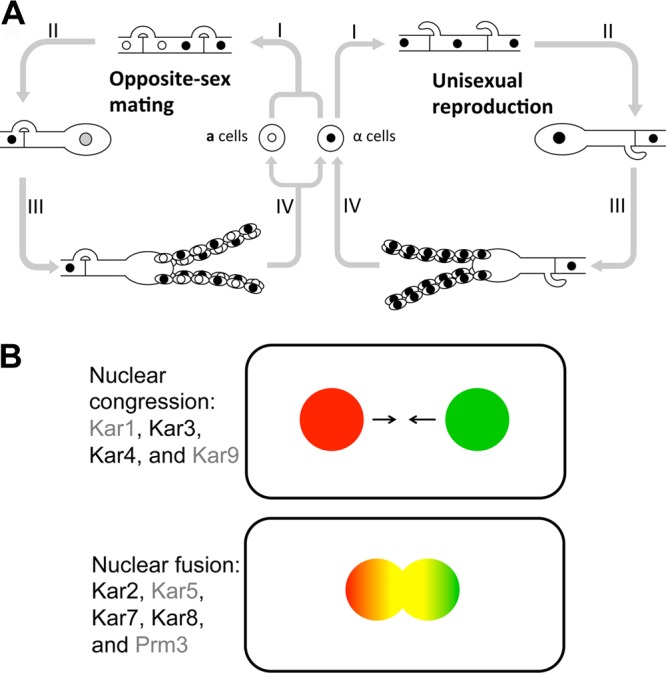
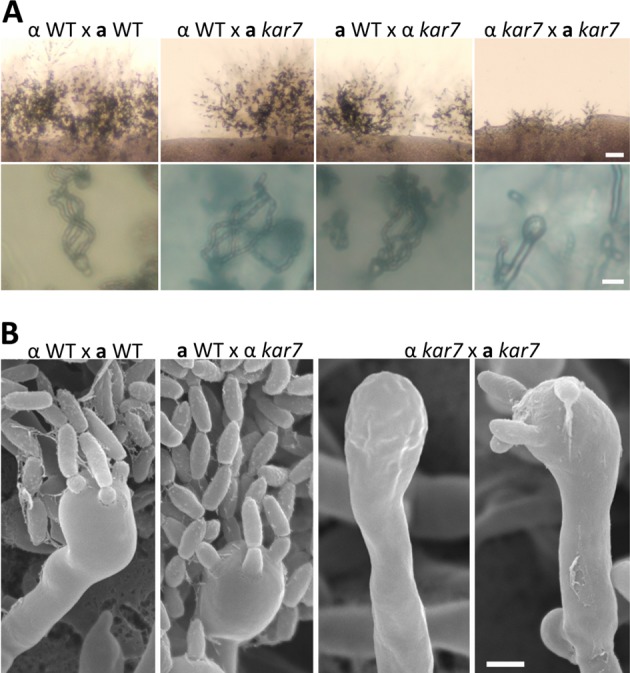
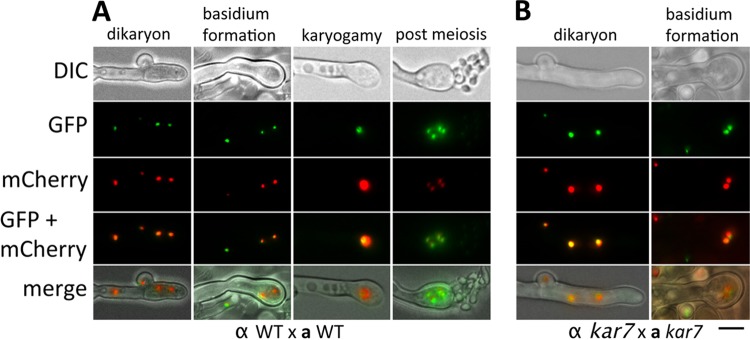
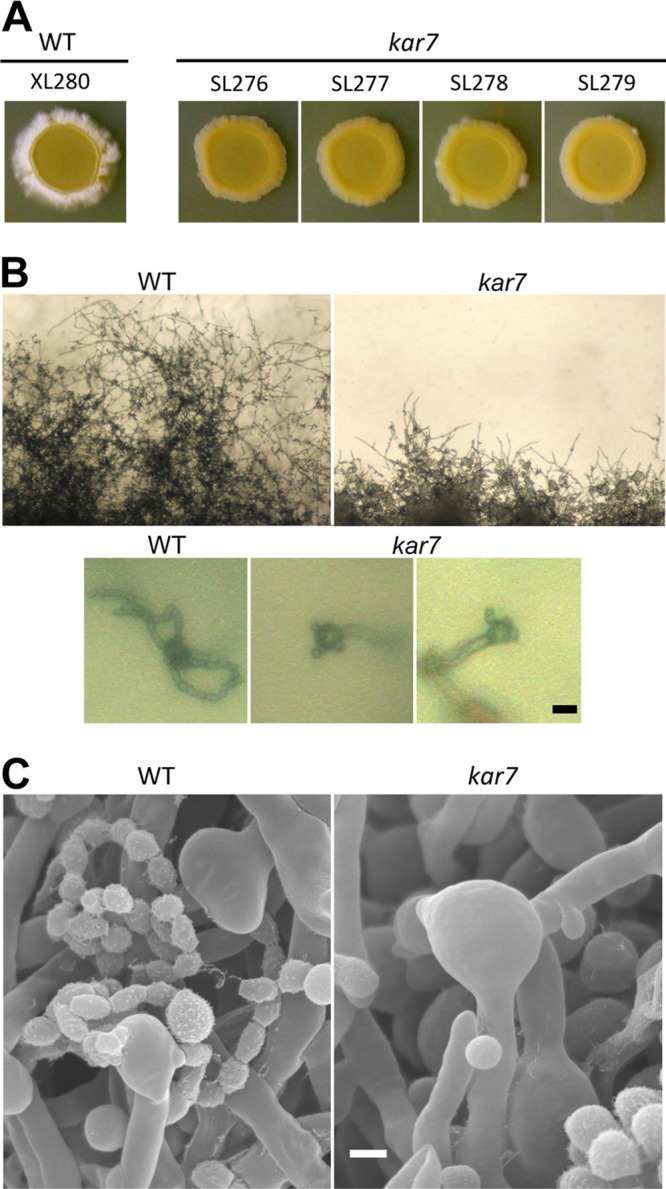
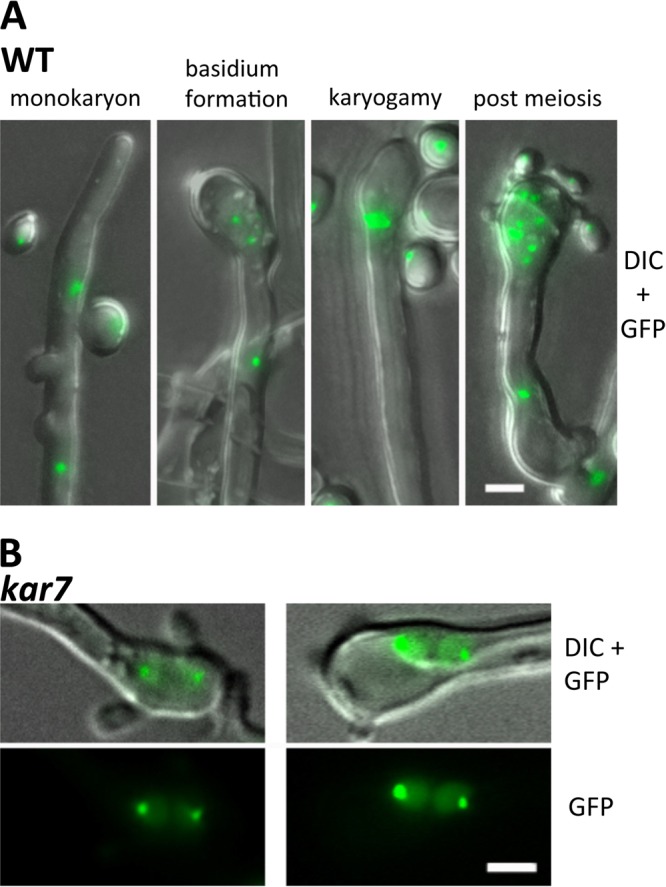
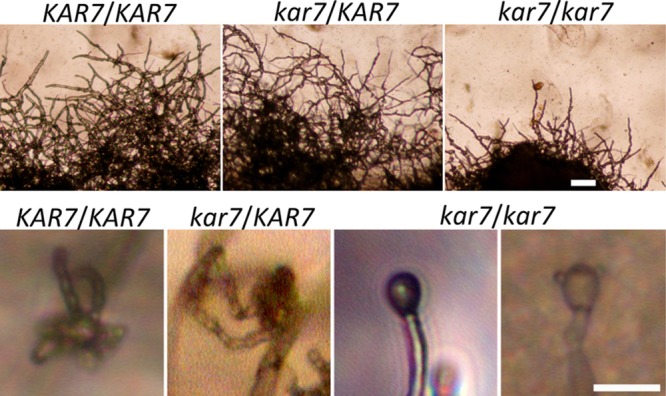
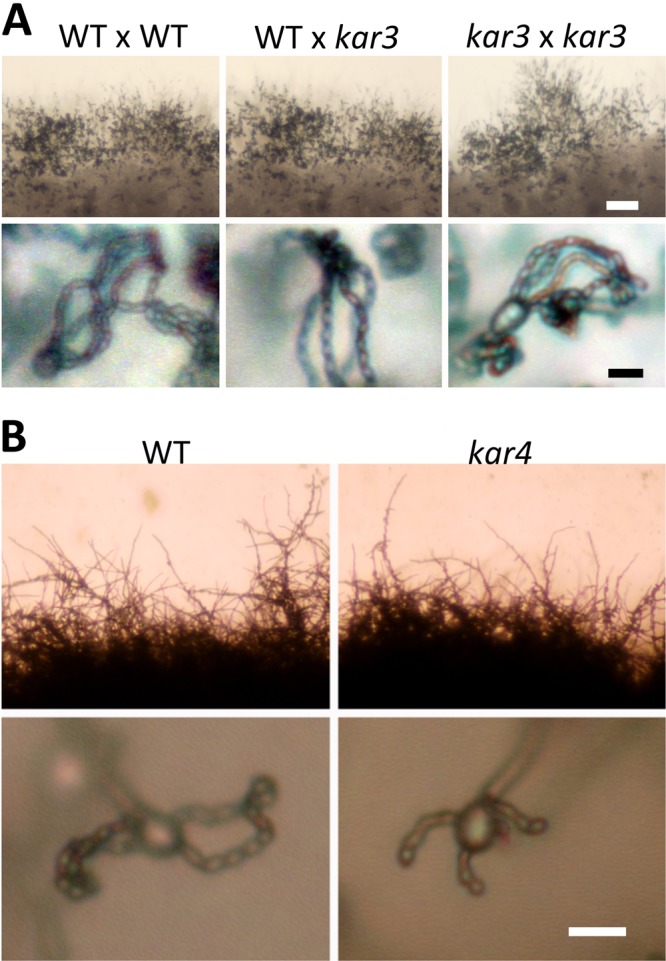
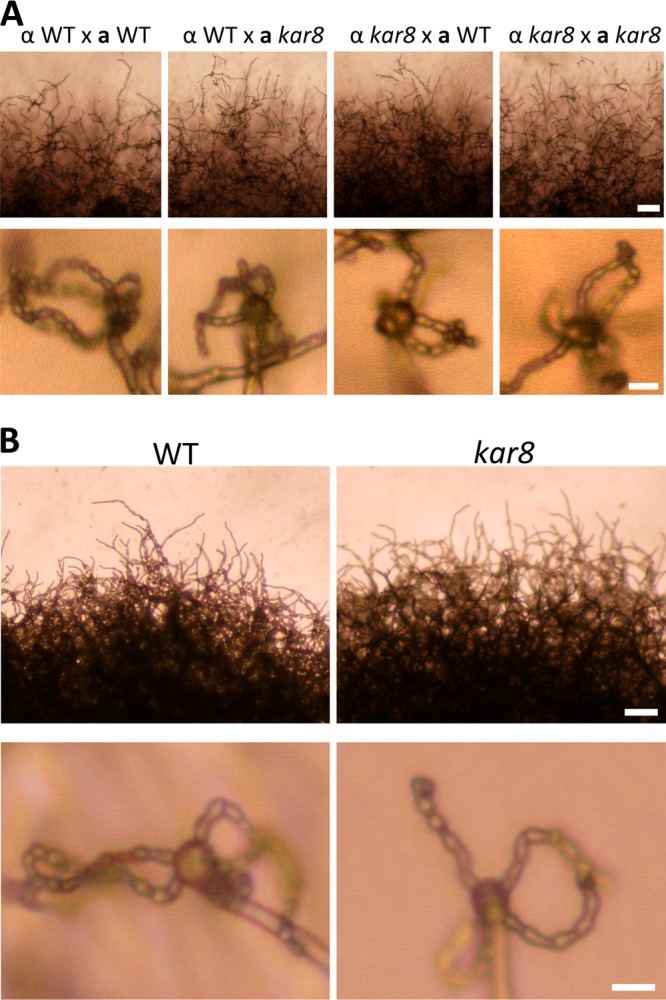
The human basidiomycetous fungal pathogen Cryptococcus neoformans serves as a model fungus to study sexual development and produces infectious propagules, basidiospores, via the sexual cycle. Karyogamy is the process of nuclear fusion and an essential step to complete mating. Therefore, regulation of nuclear fusion is central to understanding sexual development of C. neoformans. However, our knowledge of karyogamy genes was limited. In this study, using a BLAST search with the Saccharomyces cerevisiae KAR genes, we identified five C. neoformans karyogamy gene orthologs: CnKAR2, CnKAR3, CnKAR4, CnKAR7 (or CnSEC66), and CnKAR8. There are no apparent orthologs of the S. cerevisiae genes ScKAR1, ScKAR5, and ScKar9 in C. neoformans. Karyogamy involves the congression of two nuclei followed by nuclear membrane fusion, which results in diploidization. ScKar7 (or ScSec66) is known to be involved in nuclear membrane fusion. In C. neoformans, kar7 mutants display significant defects in hyphal growth and basidiospore chain formation during both a-α opposite and α-α unisexual reproduction. Fluorescent nuclear imaging revealed that during kar7 × kar7 bilateral mutant matings, the nuclei congress but fail to fuse in the basidia. These results demonstrate that the KAR7 gene plays an integral role in both opposite-sex and unisexual mating, indicating that proper control of nuclear dynamics is important. CnKAR2 was found to be essential for viability, and its function in mating is not known. No apparent phenotypes were observed during mating of kar3, kar4, or kar8 mutants, suggesting that the role of these genes may be dispensable for C. neoformans mating, which demonstrates a different evolutionary trajectory for the KAR genes in C. neoformans compared to those in S. cerevisiae.
Figures
Similar articles
-
PRM1 and KAR5 function in cell-cell fusion and karyogamy to drive distinct bisexual and unisexual cycles in the Cryptococcus pathogenic species complex.PLoS Genet. 2017 Nov 27;13(11):e1007113. doi: 10.1371/journal.pgen.1007113. eCollection 2017 Nov. PLoS Genet. 2017. PMID: 29176784 Free PMC article.
-
Uniparental nuclear inheritance following bisexual mating in fungi.Elife. 2021 Aug 2;10:e66234. doi: 10.7554/eLife.66234. Elife. 2021. PMID: 34338631 Free PMC article.
-
Unisexual reproduction enhances fungal competitiveness by promoting habitat exploration via hyphal growth and sporulation.Eukaryot Cell. 2013 Aug;12(8):1155-9. doi: 10.1128/EC.00147-13. Epub 2013 Jun 21. Eukaryot Cell. 2013. PMID: 23794511 Free PMC article.
-
[Mating types, sexual reproduction and ploidy in fungi: effects on virulence].Mikrobiyol Bul. 2009 Jul;43(3):507-13. Mikrobiyol Bul. 2009. PMID: 19795629 Review. Turkish.
-
Unisexual versus bisexual mating in Cryptococcus neoformans: Consequences and biological impacts.Fungal Genet Biol. 2015 May;78:65-75. doi: 10.1016/j.fgb.2014.08.008. Epub 2014 Aug 27. Fungal Genet Biol. 2015. PMID: 25173822 Free PMC article. Review.
Cited by
-
CryptoCEN: A Co-Expression Network for Cryptococcus neoformans reveals novel proteins involved in DNA damage repair.PLoS Genet. 2024 Feb 15;20(2):e1011158. doi: 10.1371/journal.pgen.1011158. eCollection 2024 Feb. PLoS Genet. 2024. PMID: 38359090 Free PMC article.
-
Essential roles of the Kar2/BiP molecular chaperone downstream of the UPR pathway in Cryptococcus neoformans.PLoS One. 2013;8(3):e58956. doi: 10.1371/journal.pone.0058956. Epub 2013 Mar 6. PLoS One. 2013. PMID: 23484059 Free PMC article.
-
The Evolution of Sexual Reproduction and the Mating-Type Locus: Links to Pathogenesis of Cryptococcus Human Pathogenic Fungi.Annu Rev Genet. 2019 Dec 3;53:417-444. doi: 10.1146/annurev-genet-120116-024755. Epub 2019 Sep 19. Annu Rev Genet. 2019. PMID: 31537103 Free PMC article. Review.
-
Absence of SUN-domain protein Slp1 blocks karyogamy and switches meiotic recombination and synapsis from homologs to sister chromatids.Proc Natl Acad Sci U S A. 2014 Sep 23;111(38):E4015-23. doi: 10.1073/pnas.1415758111. Epub 2014 Sep 10. Proc Natl Acad Sci U S A. 2014. PMID: 25210014 Free PMC article.
-
Sexual reproduction of human fungal pathogens.Cold Spring Harb Perspect Med. 2014 Aug 1;4(8):a019281. doi: 10.1101/cshperspect.a019281. Cold Spring Harb Perspect Med. 2014. PMID: 25085958 Free PMC article. Review.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403–410 - PubMed
-
- Anderson JB, Kohn LM. 2007. Dikaryons, diploids, and evolution, p 317–331. In Heitman J, Kronstad JW, Taylor JW, Casselton LA. (ed), Sex in fungi. ASM Press, Washington, DC
-
- Bennett RJ, Miller MG, Chua PR, Maxon ME, Johnson AD. 2005. Nuclear fusion occurs during mating in Candida albicans and is dependent on the KAR3 gene. Mol. Microbiol. 55:1046–1059 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
