

Homeodomain-interacting protein kinase (HIPK)-1 is required for splenic B cell homeostasis and optimal T-independent type 2 humoral response
- PMID: 22545114
- PMCID: PMC3335840
- DOI: 10.1371/journal.pone.0035533
Homeodomain-interacting protein kinase (HIPK)-1 is required for splenic B cell homeostasis and optimal T-independent type 2 humoral response
Abstract
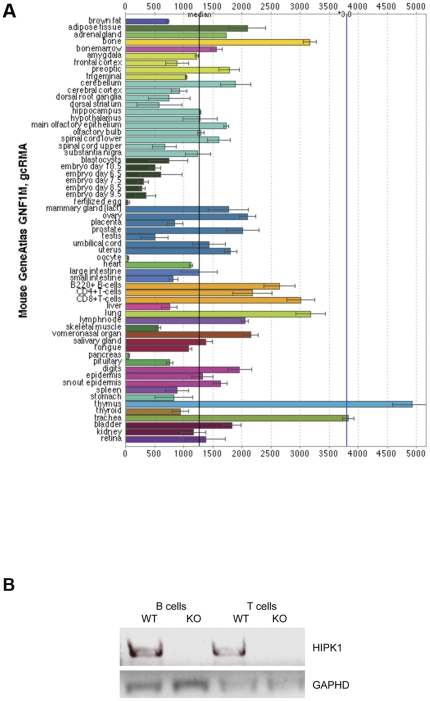
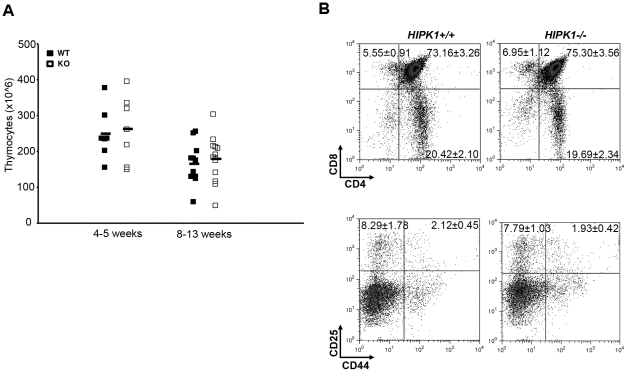
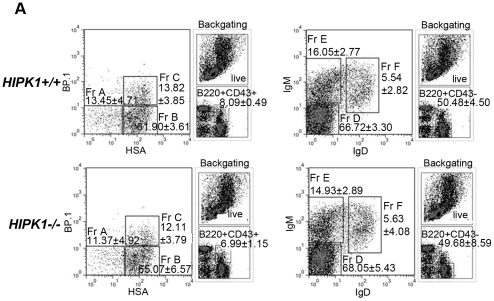
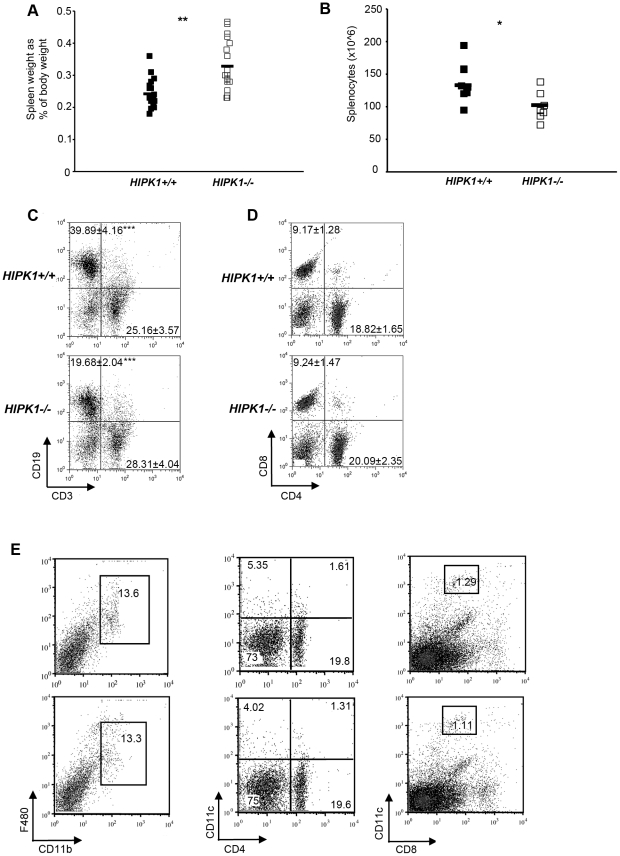
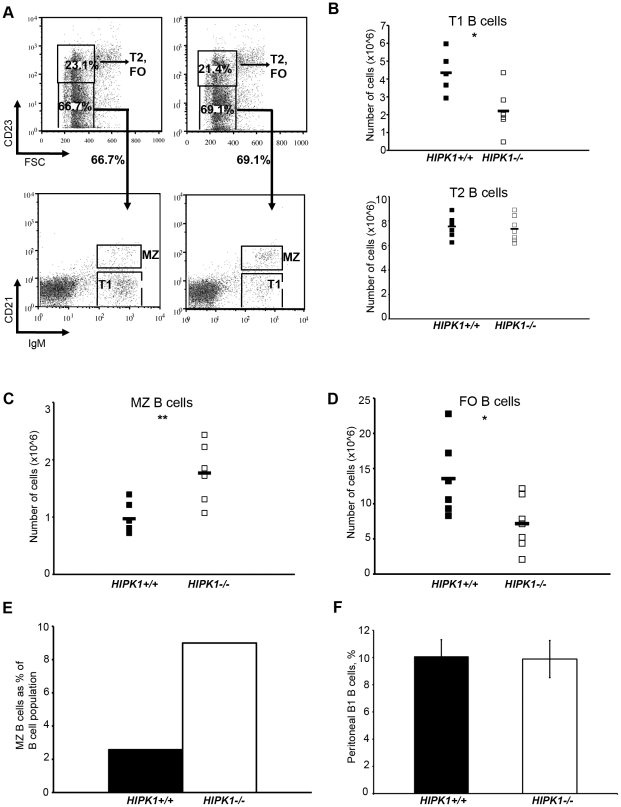
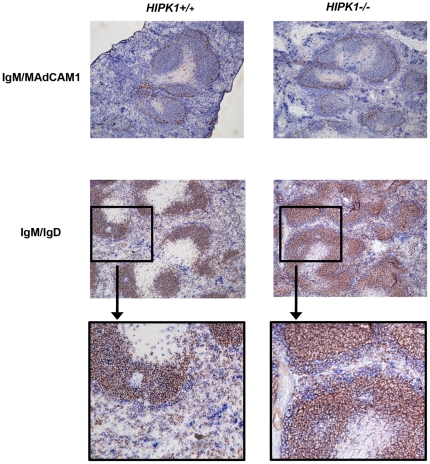
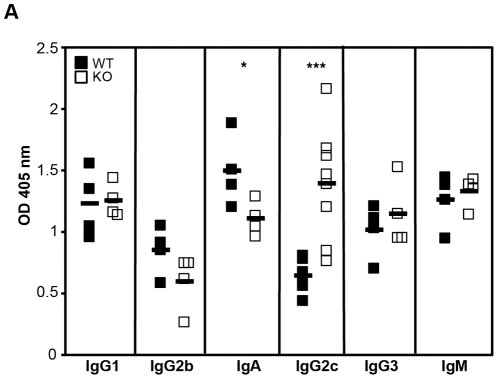
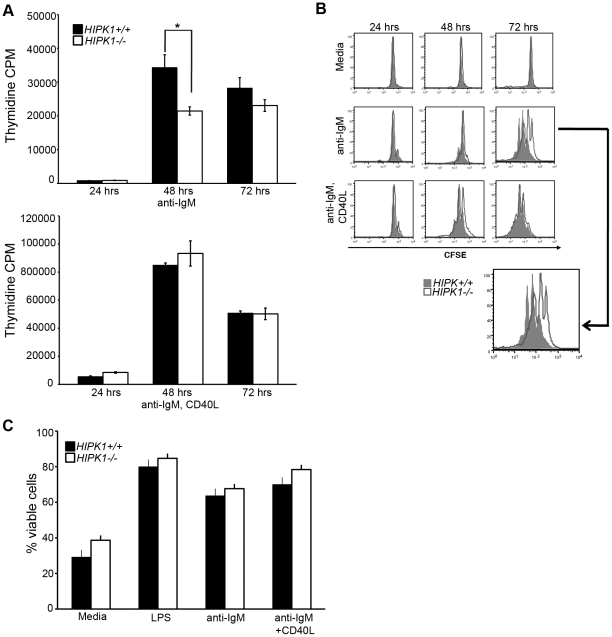
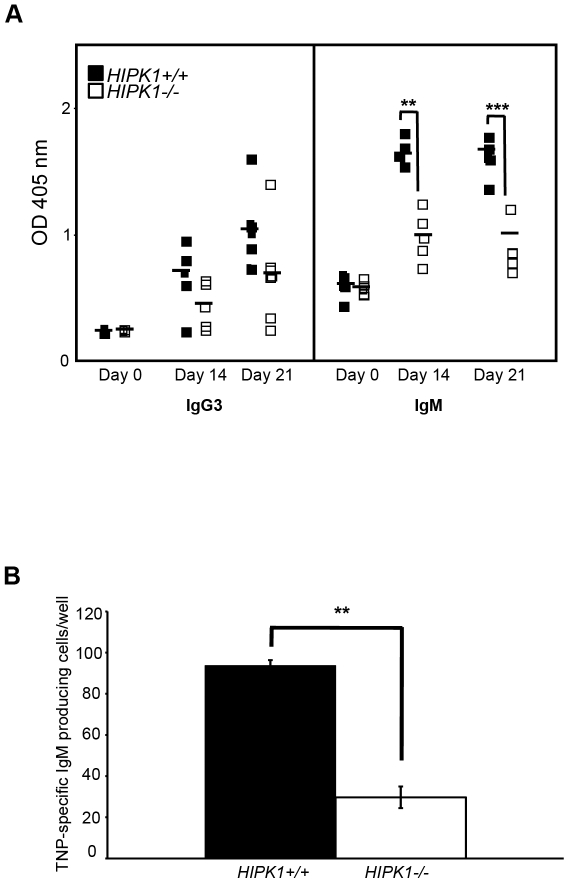
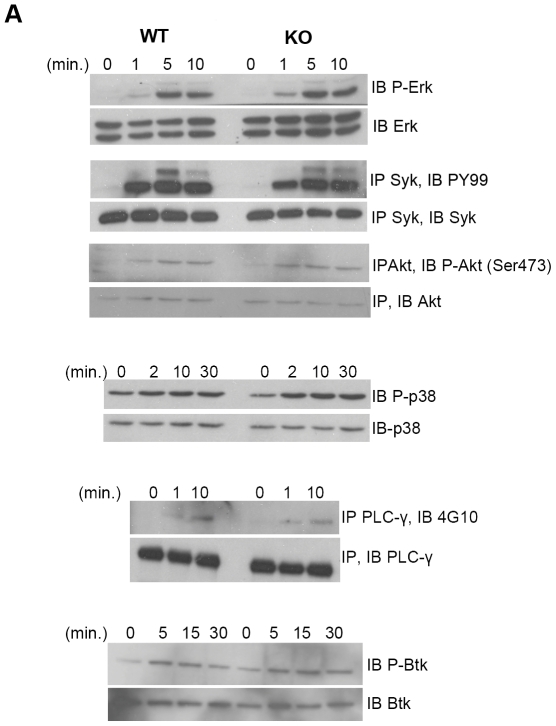
The homeodomain-interacting protein kinase (HIPK) family is comprised of four highly related serine/threonine kinases originally identified as co-repressors for various homeodomain-containing transcription factors. The HIPKs have been shown to be involved in growth regulation and apoptosis, with numerous studies highlighting HIPK regulation of the tumor suppressor p53. In this study, we have discovered a B cell homeostatic defect in HIPK1-deficient (HIPK1(-/-)) mice. Lymphopoietic populations within the thymus and bone marrow of HIPK1(-/-) mice appeared normal based upon FACS analysis; however, the spleen exhibited a reduced number of total B cells with a significant loss of transitional-1 and follicular B cell populations. Interestingly, the marginal zone B cell population was expanded in HIPK1(-/-) mice, yielding an increased frequency of these cells. HIPK1(-/-) B cells exhibited impaired cell division in response to B cell receptor cross-linking in vitro based upon thymidine incorporation or CFSE dilution; however, the addition of CD40L rescued HIPK1(-/-) proliferation to wild-type levels. Despite the expanded MZ B cell population in the HIPK1(-/-) mice, the T-independent type 2 humoral response was impaired. These data identify HIPK1 as a novel kinase required for optimal B cell function in mice.
Conflict of interest statement
Figures
Similar articles
-
Overlapping roles for homeodomain-interacting protein kinases hipk1 and hipk2 in the mediation of cell growth in response to morphogenetic and genotoxic signals.Mol Cell Biol. 2006 Apr;26(7):2758-71. doi: 10.1128/MCB.26.7.2758-2771.2006. Mol Cell Biol. 2006. PMID: 16537918 Free PMC article.
-
Characterization of cells and gene-targeted mice deficient for the p53-binding kinase homeodomain-interacting protein kinase 1 (HIPK1).Proc Natl Acad Sci U S A. 2003 Apr 29;100(9):5431-6. doi: 10.1073/pnas.0530308100. Epub 2003 Apr 17. Proc Natl Acad Sci U S A. 2003. PMID: 12702766 Free PMC article.
-
Homeodomain-interacting protein kinase 1 modulates Daxx localization, phosphorylation, and transcriptional activity.Mol Cell Biol. 2003 Feb;23(3):950-60. doi: 10.1128/MCB.23.3.950-960.2003. Mol Cell Biol. 2003. PMID: 12529400 Free PMC article.
-
Update on the Regulation of HIPK1, HIPK2 and HIPK3 Protein Kinases by microRNAs.Microrna. 2018;7(3):178-186. doi: 10.2174/2211536607666180525102330. Microrna. 2018. PMID: 29793420 Review.
-
Integration of stress signals by homeodomain interacting protein kinases.Biol Chem. 2014 Apr;395(4):375-86. doi: 10.1515/hsz-2013-0264. Biol Chem. 2014. PMID: 24225127 Review.
Cited by
-
Cell extrinsic alterations in splenic B cell maturation in Flt3-ligand knockout mice.Immun Inflamm Dis. 2015 Jun;3(2):103-17. doi: 10.1002/iid3.54. Epub 2015 Apr 15. Immun Inflamm Dis. 2015. PMID: 26029370 Free PMC article.
-
Mapping effector genes at lupus GWAS loci using promoter Capture-C in follicular helper T cells.Nat Commun. 2020 Jul 3;11(1):3294. doi: 10.1038/s41467-020-17089-5. Nat Commun. 2020. PMID: 32620744 Free PMC article.
-
Phosphorylation of Krüppel-like factor 3 (KLF3/BKLF) and C-terminal binding protein 2 (CtBP2) by homeodomain-interacting protein kinase 2 (HIPK2) modulates KLF3 DNA binding and activity.J Biol Chem. 2015 Mar 27;290(13):8591-605. doi: 10.1074/jbc.M115.638338. Epub 2015 Feb 6. J Biol Chem. 2015. PMID: 25659434 Free PMC article.
-
Identification of ROCK1 as a novel biomarker for postmenopausal osteoporosis and pan-cancer analysis.Aging (Albany NY). 2023 Sep 7;15(17):8873-8907. doi: 10.18632/aging.205004. Epub 2023 Sep 7. Aging (Albany NY). 2023. PMID: 37683138 Free PMC article.
-
Coordinated signals from the DNA repair enzymes PARP-1 and PARP-2 promotes B-cell development and function.Cell Death Differ. 2019 Dec;26(12):2667-2681. doi: 10.1038/s41418-019-0326-5. Epub 2019 Apr 17. Cell Death Differ. 2019. PMID: 30996287 Free PMC article.
References
-
- Oliver AM, Martin F, Gartland GL, Carter RH, Kearney JF. Marginal zone B cells exhibit unique activation, proliferative and immunoglobulin secretory responses. Eur J Immunol. 1997;27(9):2366–2374. - PubMed
-
- Martin F, Kearney JF. Marginal-zone B cells. Nat Rev Immunol. 2002;2(5):323–335. - PubMed
-
- Cariappa A, Mazo IB, Chase C, Shi HN, Liu H, et al. Perisinusoidal B cells in the bone marrow participate in T-independent responses to blood-borne microbes. Immunity. 2005;23(4):397–407. - PubMed
-
- Cariappa A, Chase C, Liu H, Russell P, Pillai S. Naive recirculating B cells mature simultaneously in the spleen and bone marrow. Blood. 2007;109(6):2339–2345. - PubMed
-
- Balazs M, Martin F, Zhou T, Kearney J. Blood dendritic cells interact with splenic marginal zone B cells to initiate T-independent immune responses. Immunity. 2002;17(3):341–352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
