

doi: 10.4049/jimmunol.1102629.
Epub 2012 Apr 30.
Altered Ig hypermutation pattern and frequency in complementary mouse models of DNA polymerase ζ activity
Affiliations
- PMID: 22547703
- PMCID: PMC3358437
- DOI: 10.4049/jimmunol.1102629
Item in Clipboard
Altered Ig hypermutation pattern and frequency in complementary mouse models of DNA polymerase ζ activity
J Immunol.
.
Abstract
To test the hypothesis that DNA polymerase ζ participates in Ig hypermutation, we generated two mouse models of Pol ζ function: a B cell-specific conditional knockout and a knock-in strain with a Pol ζ mutagenesis-enhancing mutation. Pol ζ-deficient B cells had a reduction in mutation frequency at Ig loci in the spleen and in Peyer's patches, whereas knock-in mice with a mutagenic Pol ζ displayed a marked increase in mutation frequency in Peyer's patches, revealing a pattern that was similar to mutations in yeast strains with a homologous mutation in the gene encoding the catalytic subunit of Pol ζ. Combined, these data are best explained by a direct role for DNA polymerase ζ in Ig hypermutation.
Figures

Normal B cell development in Rev3 knock-out and Rev3L2610Fmice. (A) B cells from 8–12 week-old mice were sorted and examined by FACS. Development of Rev3 knock-out and Rev3L2610FB cells in bone marrow stage pre-BII (left (B220+ CD25+) and in the immature B cell stage (middle, B220+ IgM+) is comparable to CD19-promoter driven cre-recombinase transgenics (heterogyzotes) and C57BL6 controls (not shown). As the B cells mature and migrate to the spleen (right, B220+IgM+) normal numbers of mature/resting splenic B cell populations are observed in both the Rev3 knock-out and Rev3L2610F. (B) Unimmunized12 week-old mice were NP-CGG injected and their B cells examined 15 days later. The activated splenic GC B cell population in Rev3 knock-out mice undergoing somatic hypermutation (B220+GL7+) appears normal in numbers with 42–56% of this gated population being YFP+. However, while the percentage of GL7+ B cells in PP appears normal, only a tiny fraction of those (3%) were YFP+. (C) Enrichment for activated splenic B cells from conditional Rev3 knock-out, 15 days after immunization with 150µg NP-CGG in alum. The post-sort (B220+ YFP+ λ+) GC splenic B cell population is greatly enriched for the YFP (marker for Rev3 deletion). The λ light chain was selected because it is associated with the Vh186.2 HC for the high affinity response to the NP hapten. This pure Rev3-null population was used for subsequent RTPCR and mutation analysis (D) Over 90% of B220+ B cells in the spleen were YFP+ in Rev3 knock-out mice.
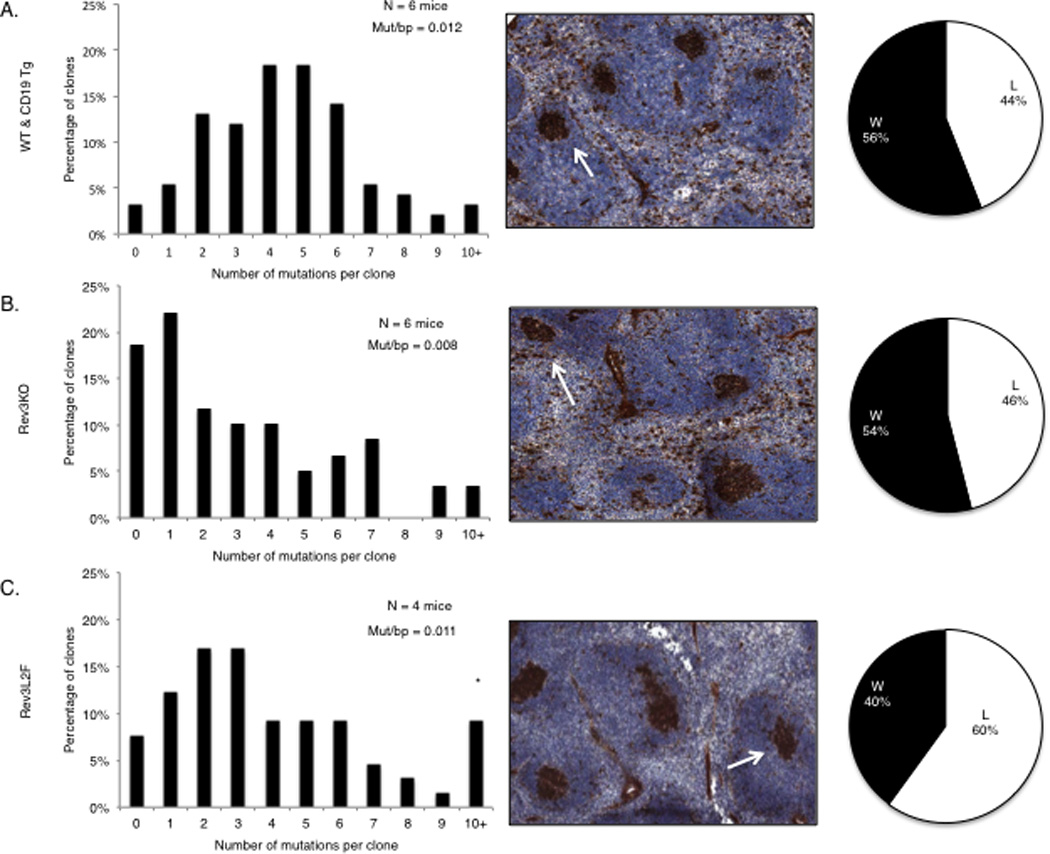
The immune response in mice with altered DNA polymerase ζ function. (A–B) Rev3 knock-out B cells experienced a decrease in the frequency of mutations at Ig V regions during the immune response to NP-CGG compared to WT mice and CD19 transgenics. Close to 40% of the clones in the Rev3 knock-out had 0–1 mutations, compared to less than 10% in the controls. Spleens from Rev3 knock-out mice displayed normal GC with 46% of the clones achieving the affinity-enhancing W to L mutation at position 33 of Vh186.2. (C) Rev3 knock-in mice (Rev3L2610F) had a similar overall distribution of mutational classes as controls but had a higher percentage of clones with more than 9 mutations. These mice displayed normal GC, but also displayed an increase in the fraction of Vh186.2 clones with the affinity enhancing W to L change (pie chart). The following mice were used for these experiments: C57BL/6 and CD19 promoter-driven cre recombinase transgenics in the C57BL/6 background, n = 6 mice and 75 unique clones; Rev3 knock-out, n = 6 mice and 59 unique clones; Rev3L2610Fclones, n = 4 mice and 50 unique clones. Unique clones were determined by the CDR3 amino acid sequence.
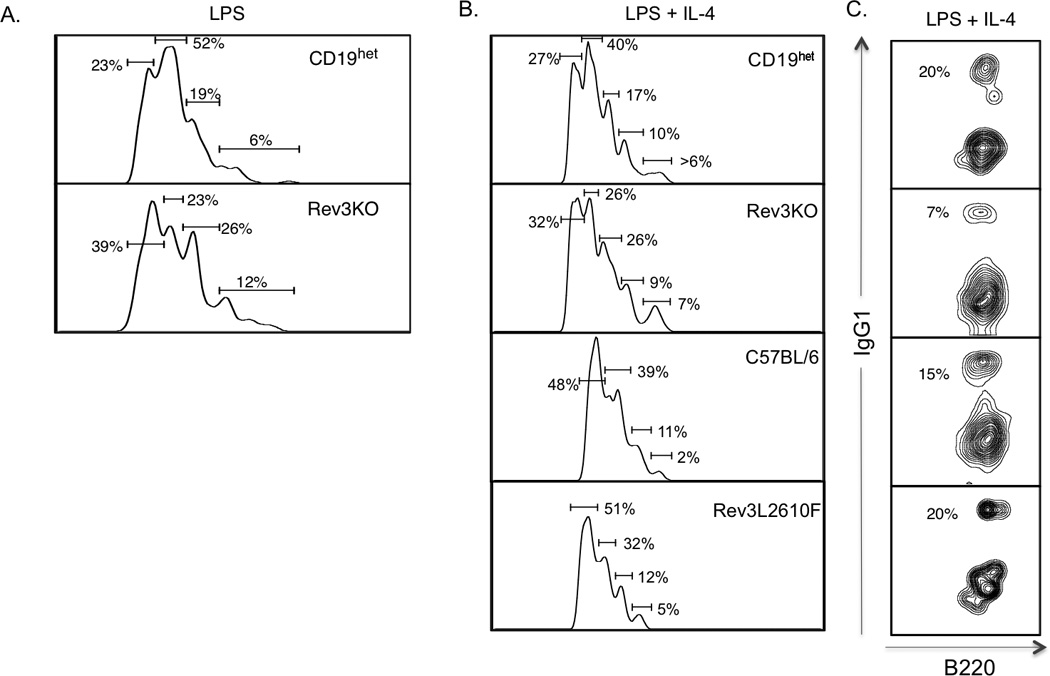
B cell proliferation in Rev3KO and Rev3L2610F mice. CD43-depleted splenic B cells from 8–12 week-old mice were isolated and incubated with 5nM CFSE in PBS to measure cell proliferation. B cells (106 cells/ml) were stimulated over 4 days using LPS alone (20ug/ml) (A), or together with IL-4 (25ng/ml) (B) to stimulate class switch recombination. In vitro stimulation of B cells was determined at day 0 and day 4 of stimulation. Flow cytometry was used to determine the B cell proliferation following in vitro stimulation as depicted by histograms. The numbered gated represent the percentage of B cells within sequential rounds of cell division. These proliferation peaks were based on 105 events, duplicate samples and a total of 5 mice per group. (C) Percentage of IgG1+ B cells following 4 days of activation with LPS and IL4.
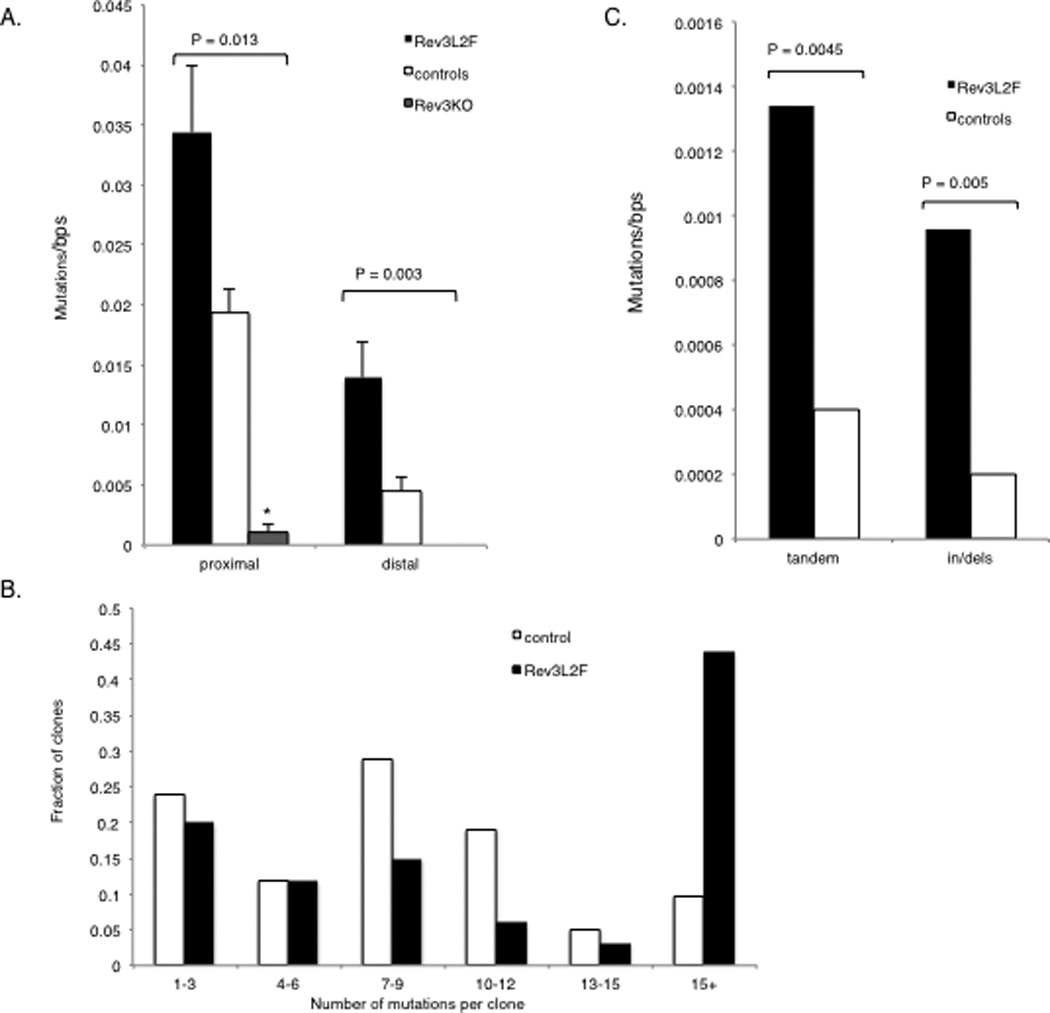
Mutations at the intronic region downstream of rearranged V genes in Peyer’s Patches B cells in mice with altered DNA polymerase ζ function. The IgH intronic region downstream of rearranged V genes of GC B cells was assessed for accumulation of mutations. Peyer’s Patches B220+ GL7+ (and YFP+ for Rev3 knock-out) B lymphocytes were isolated from mice aged 12 or 36 weeks: C57BL/6, n = 20 mice and 76 unique clones (with 53% of the mutated clones from 36 weeks old mice); Rev3L2610Fn = 8 mice and 84 unique clones (with 58% of the mutated clones from 36 weeks old mice); Rev3 knock-out, n = 4 mice and 18 unique clones (with 100% of the clones from 36 weeks old mice). Because the rearranged CDR3 was included in the PCR fragment, unique clones could be assessed and those differing by more than 5 mutations were considered as different clones, although shared mutations were only counted once. (A) Mutations per nucleotide among mutated clones were calculated for both the proximal (~ 430 bps, beginning immediately downstream of JH4) and distal ends of the IgH intronic region downstream of rearranged V genes (region was the last 590 bps of the 1.2kb PCR fragment). The regions were analyzed separately to examine mutation pattern and frequency in less saturated regions. In both the proximal and distal regions, the mutation rate in the Rev3L2610Fmice is at least twice that observed in controls and this difference was highly significant. Rev3 knock-out clones had an extremely low mutation frequency that was highly significantly different from controls. All except one of the unique Rev3 knock-out clones had no mutation; thus, all were included in the analysis. (B) The increase in mutation frequency among mutated clones in Rev3L2610Fmice was the result of clones with a large number of mutations: there was a dramatic increase in the fraction of Rev3L2610Fclones with >15 mutations, some having as many as 30 or more mutations in the proximal region alone (P = 0.037). 98% of the Rev3 knock-out clones were unmutated even in 6 month-old mice, compared to 33 and 57% for the control and the Rev3L2610Fclones, respectively. (C) Tandem doublets and triplets and insertions and deletions were significantly increased in Rev3L2610Fmice. The difference in mutation frequencies were tested for significance using the Mann-Whitney U test, the difference in the distribution of mutations among clones was tested for significance using the Kolmogorov-Smirnov test and the difference in the frequency of tandem and indels among all mutations was analyzed using the Fisher exact test (two-sided p values are given).
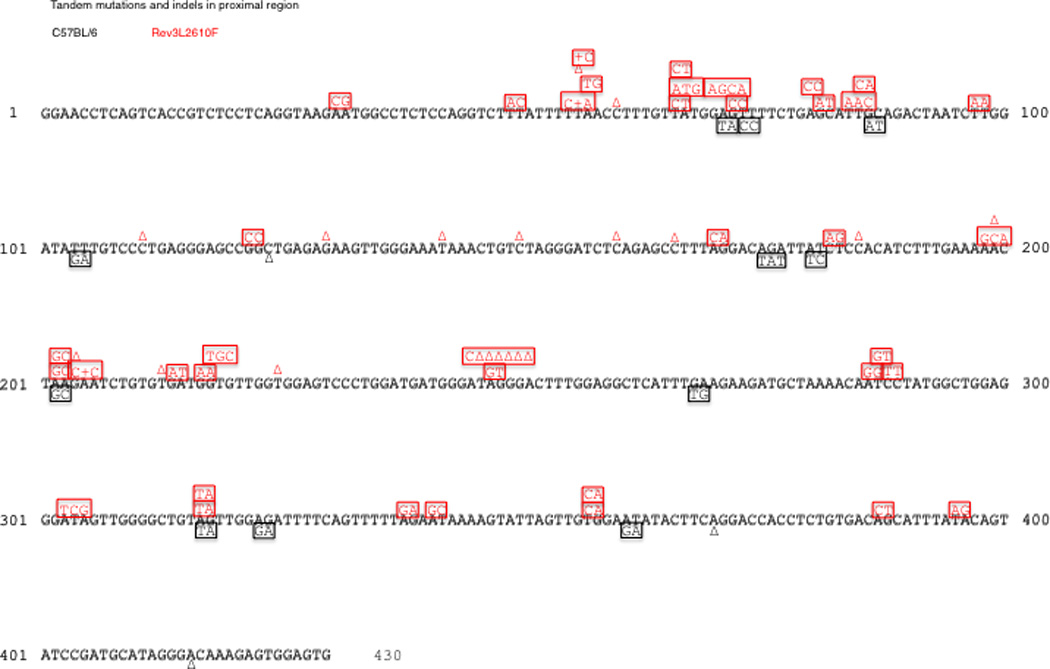
Tandem and indel mutations in the proximal HC intronic region. B cells from Rev3L2610F mice (33 unique clones) reveal an increase in tandem (squares) and indel mutations (+ or Δ) when compared to controls (41 unique clones). A similar pattern is seen in the distal region of the sequences (data not shown). Shared mutations from oligoclonality and single base substitutions are not shown.
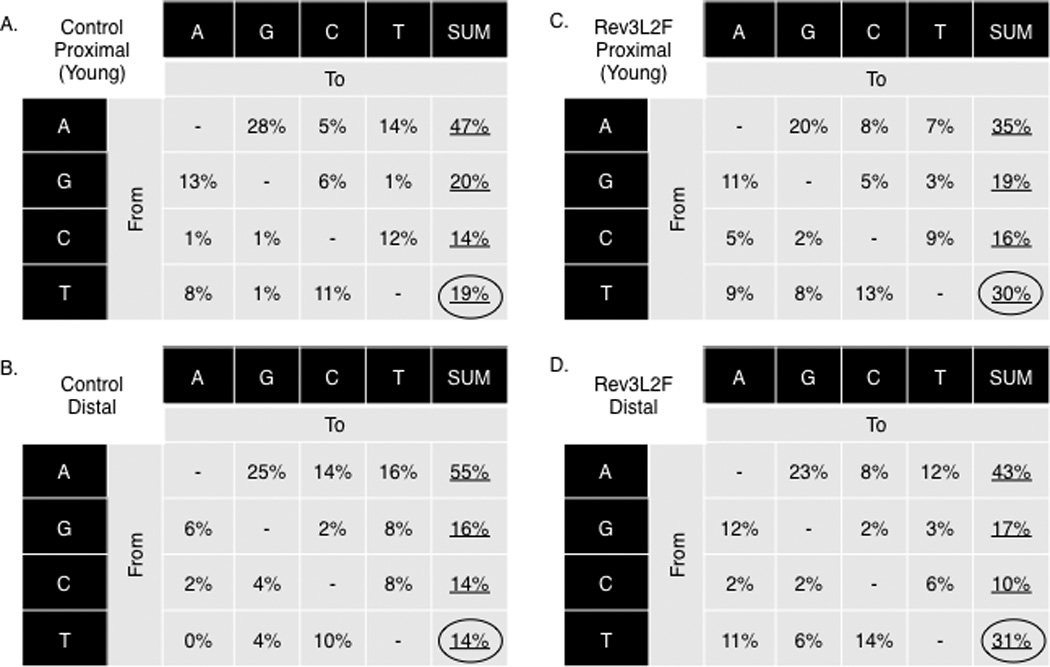
Mutational pattern of Rev3L2610Fmice reveals a moderate increase in mutations at T bases indicating Rev3L2610F-mediated strand bias. The mutation pattern at the IgH intronic proximal regions of B cells from young mice is depicted for controls (A) and Rev3L2610F(B) mice. The IgH intronic distal region of all controls (C) and all Rev3L2610F(D) mice B cells are also shown. The combined increase at thymidine was statistically significant (χ2 test, P = 0.04). The base composition in the proximal region is 57% A: T and 43% G:C, and for the distal region, it is 61% A:T and 39% G:C.
References
-
- Weigert MG, Cesari IM, Yonkovich SJ, Cohn M. Variability in the lambda light chain sequences of mouse antibody. Nature. 1970;228:1045–1047. - PubMed
-
- Muramatsu M, Kinoshita K, Fagarasan S, Yamada S, Shinkai Y, Honjo T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 2000;102:553–563. - PubMed
-
- Revy P, Muto T, Levy Y, Geissmann F, Plebani A, Sanal O, Catalan N, Forveille M, Dufourcq-Labelouse R, Gennery A, Tezcan I, Ersoy F, Kayserili H, Ugazio AG, Brousse N, Muramatsu M, Notarangelo LD, Kinoshita K, Honjo T, Fischer A, Durandy A. Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the Hyper-IgM syndrome (HIGM2) Cell. 2000;102:565–575. - PubMed
-
- Petersen-Mahrt SK, Harris RS, Neuberger MS. AID mutates E. coli suggesting a DNA deamination mechanism for antibody diversification. Nature. 2002;418:99–103. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
