

Mycobacterium tuberculosis Eis protein initiates suppression of host immune responses by acetylation of DUSP16/MKP-7
- PMID: 22547814
- PMCID: PMC3356628
- DOI: 10.1073/pnas.1120251109
Mycobacterium tuberculosis Eis protein initiates suppression of host immune responses by acetylation of DUSP16/MKP-7
Abstract
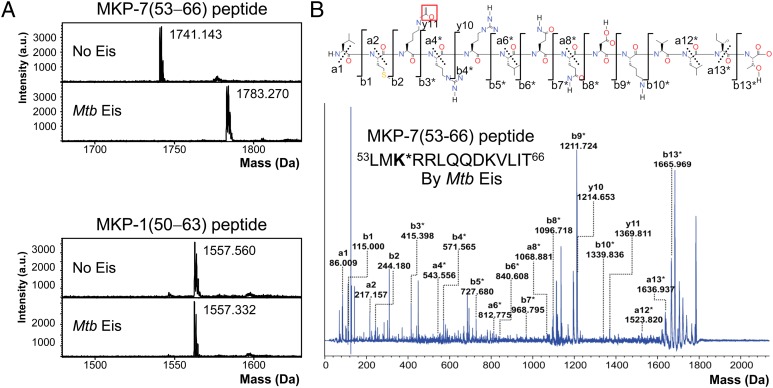
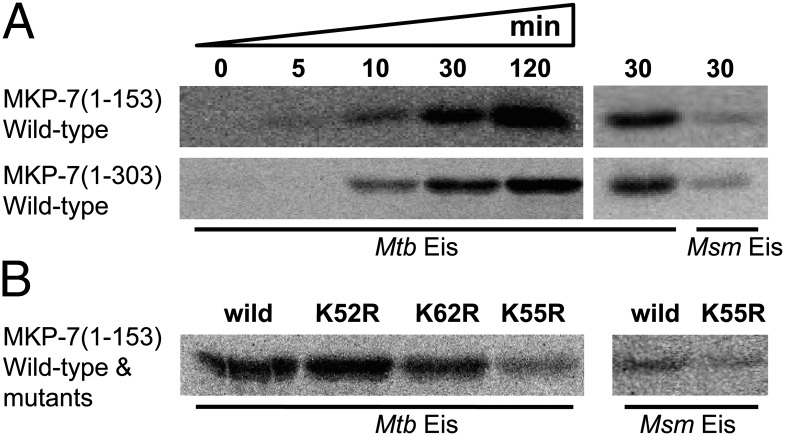
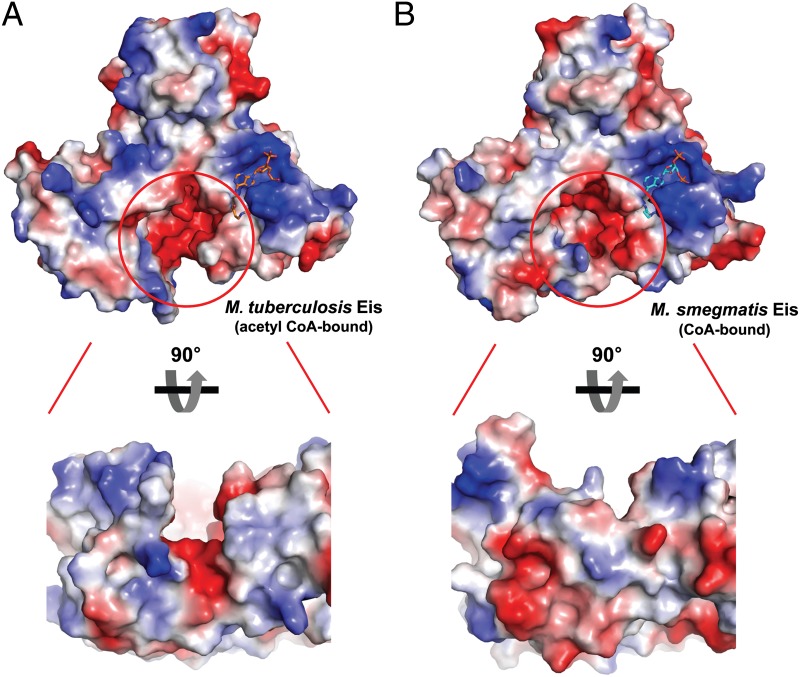
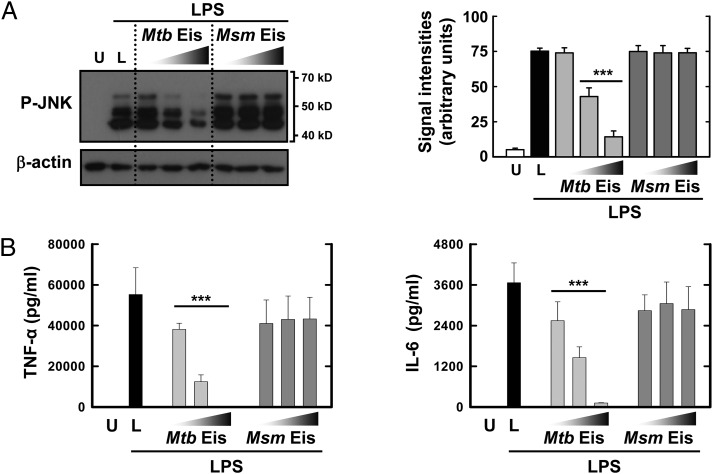
The intracellular pathogen Mycobacterium tuberculosis (Mtb) causes tuberculosis. Enhanced intracellular survival (Eis) protein, secreted by Mtb, enhances survival of Mycobacterium smegmatis (Msm) in macrophages. Mtb Eis was shown to suppress host immune defenses by negatively modulating autophagy, inflammation, and cell death through JNK-dependent inhibition of reactive oxygen species (ROS) generation. Mtb Eis was recently demonstrated to contribute to drug resistance by acetylating multiple amines of aminoglycosides. However, the mechanism of enhanced intracellular survival by Mtb Eis remains unanswered. Therefore, we have characterized both Mtb and Msm Eis proteins biochemically and structurally. We have discovered that Mtb Eis is an efficient N(ε)-acetyltransferase, rapidly acetylating Lys55 of dual-specificity protein phosphatase 16 (DUSP16)/mitogen-activated protein kinase phosphatase-7 (MKP-7), a JNK-specific phosphatase. In contrast, Msm Eis is more efficient as an N(α)-acetyltransferase. We also show that Msm Eis acetylates aminoglycosides as readily as Mtb Eis. Furthermore, Mtb Eis, but not Msm Eis, inhibits LPS-induced JNK phosphorylation. This functional difference against DUSP16/MKP-7 can be understood by comparing the structures of two Eis proteins. The active site of Mtb Eis with a narrow channel seems more suitable for sequence-specific recognition of the protein substrate than the pocket-shaped active site of Msm Eis. We propose that Mtb Eis initiates the inhibition of JNK-dependent autophagy, phagosome maturation, and ROS generation by acetylating DUSP16/MKP-7. Our work thus provides insight into the mechanism of suppressing host immune responses and enhancing mycobacterial survival within macrophages by Mtb Eis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dye C, Williams BG. The population dynamics and control of tuberculosis. Science. 2010;328:856–861. - PubMed
-
- Chiang CY, Centis R, Migliori GB. Drug-resistant tuberculosis: Past, present, future. Respirology. 2010;15:413–432. - PubMed
-
- Getahun H, Gunneberg C, Granich R, Nunn P. HIV infection-associated tuberculosis: The epidemiology and the response. Clin Infect Dis. 2010;50(Suppl 3):S201–S207. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
