

Evidence for the importance of OxPAPC interaction with cysteines in regulating endothelial cell function
- PMID: 22550136
- PMCID: PMC3371242
- DOI: 10.1194/jlr.M025320
Evidence for the importance of OxPAPC interaction with cysteines in regulating endothelial cell function
Abstract
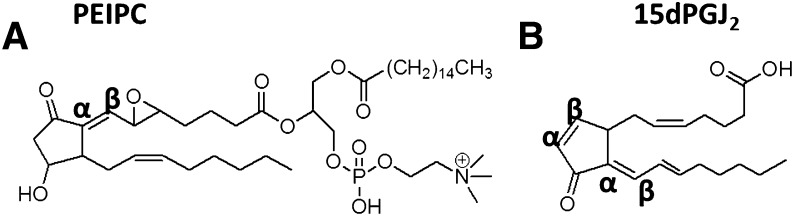
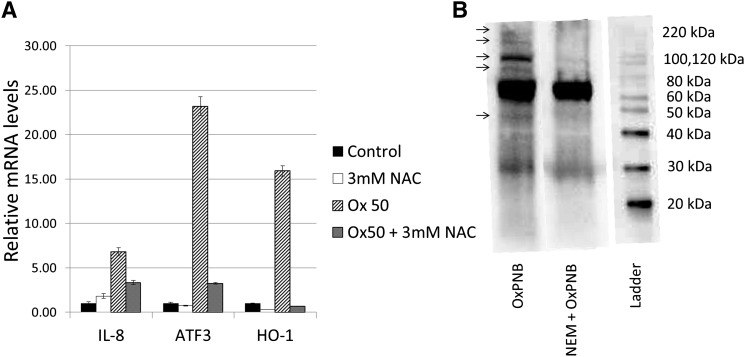
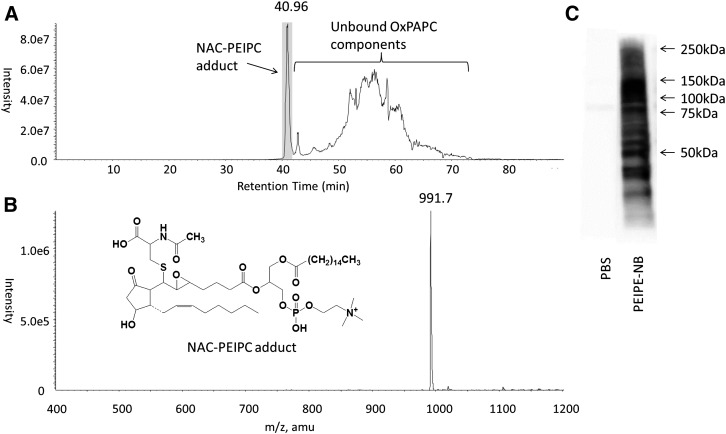
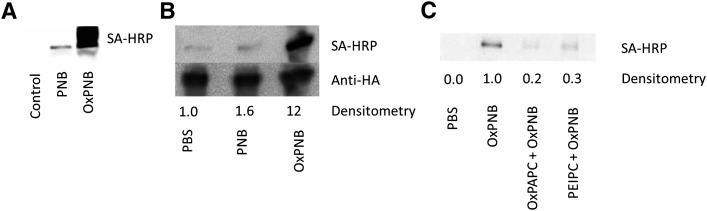
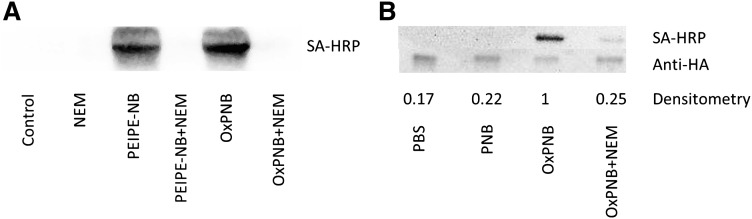
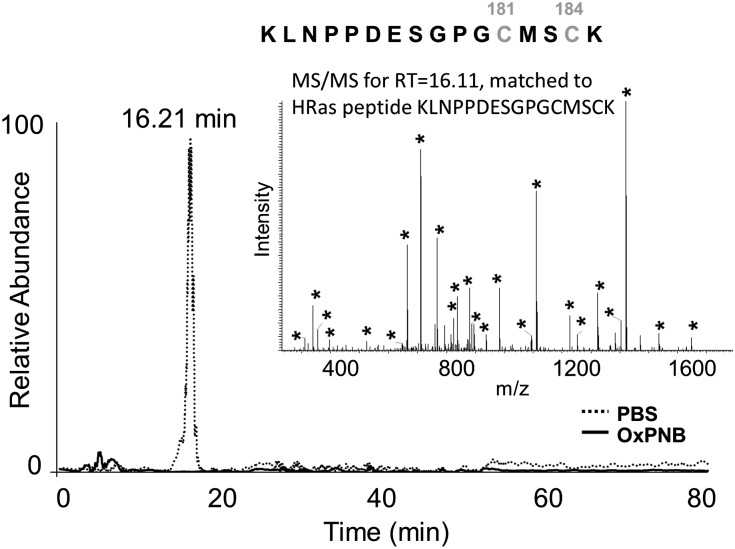
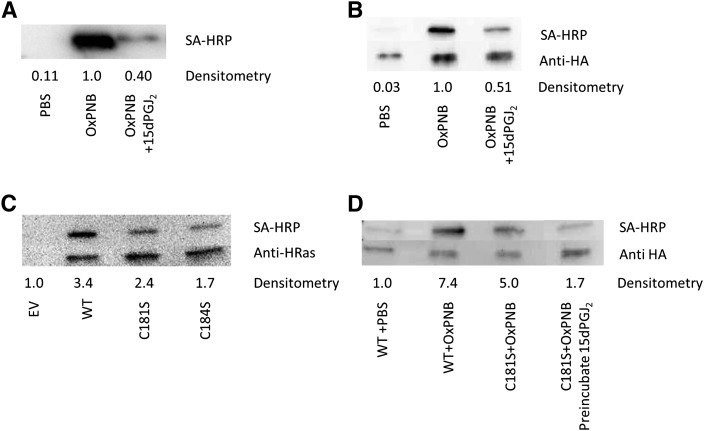
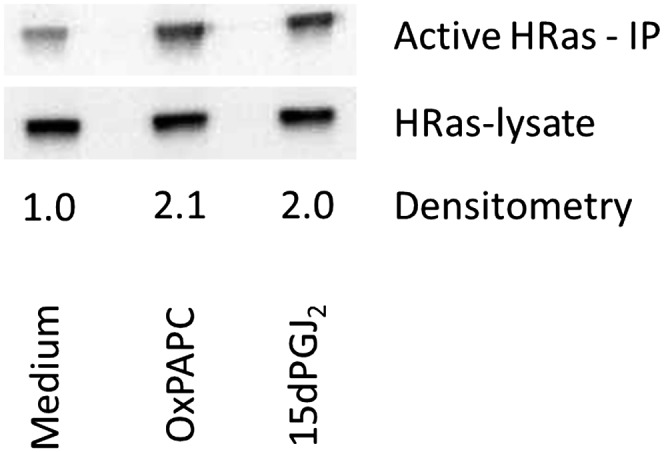
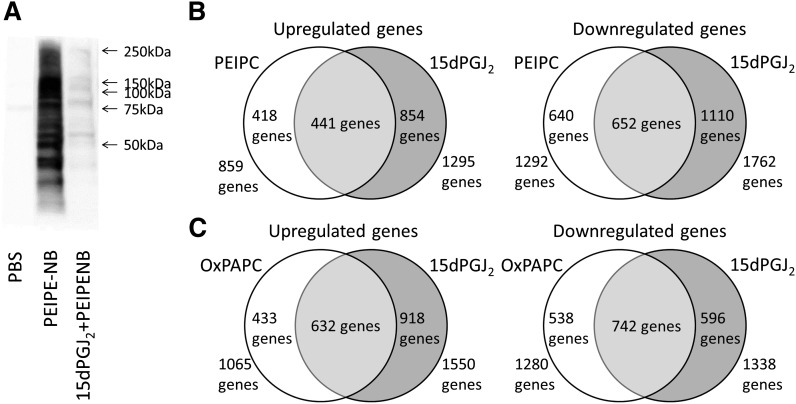
Oxidation products of 1-palmitoyl-2-arachidonoyl-sn-glycerol-3-phosphatidylcholine (PAPC), referred to as OxPAPC, and an active component, 1-palmitoyl-2-(5,6-epoxyisoprostane E₂)-sn-glycero-3-phosphatidylcholine (PEIPC), accumulate in atherosclerotic lesions and regulate over 1,000 genes in human aortic endothelial cells (HAEC). We previously demonstrated that OxPNB, a biotinylated analog of OxPAPC, covalently binds to a number of proteins in HAEC. The goal of these studies was to gain insight into the binding mechanism and determine whether binding regulates activity. In whole cells, N-acetylcysteine inhibited gene regulation by OxPAPC, and blocking cell cysteines with N-ethylmaleimide strongly inhibited the binding of OxPNB to HAEC proteins. Using MS, we demonstrate that most of the binding of OxPAPC to cysteine is mediated by PEIPC. We also show that OxPNB and PEIPE-NB, the analog of PEIPC, bound to a model protein, H-Ras, at cysteines previously shown to regulate activity in response to 15-deoxy-Δ12,14-prostaglandin J2 (15dPGJ₂). This binding was observed with recombinant protein and in cells overexpressing H-Ras. OxPAPC and PEIPC compete with OxPNB for binding to H-Ras. 15dPGJ₂ and OxPAPC increased H-Ras activity at comparable concentrations. Using microarray analysis, we demonstrate a considerable overlap of gene regulation by OxPAPC, PEIPC, and 15dPGJ₂ in HAEC, suggesting that some effects attributed to 15dPGJ₂ may also be regulated by PEIPC because both molecules accumulate in inflammatory sites. Overall, we provide evidence for the importance of OxPAPC-cysteine interactions in regulating HAEC function.
Figures
References
-
- Berliner J. A., Navab M., Fogelman A. M., Frank J. S., Demer L. L., Edwards P. A., Watson A. D., Lusis A. J. 1995. Atherosclerosis: basic mechanisms. oxidation, inflammation, and genetics. Circulation. 91: 2488–2496 - PubMed
-
- Gugiu B. G., Mouillesseaux K., Duong V., Herzog T., Hekimian A., Koroniak L., Vondriska T. M., Watson A. D. 2008. Protein targets of oxidized phospholipids in endothelial cells. J. Lipid Res. 49: 510–520 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
