

Mammalian kidney development: principles, progress, and projections
- PMID: 22550230
- PMCID: PMC3331696
- DOI: 10.1101/cshperspect.a008300
Mammalian kidney development: principles, progress, and projections
Abstract
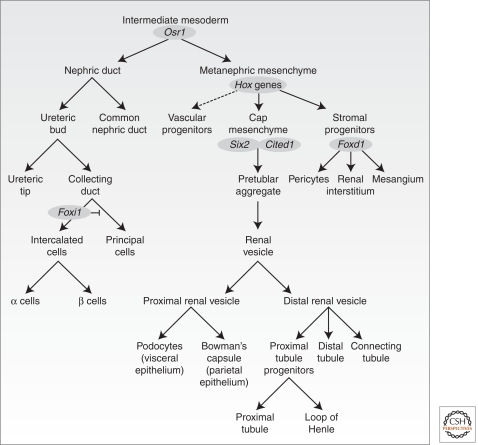
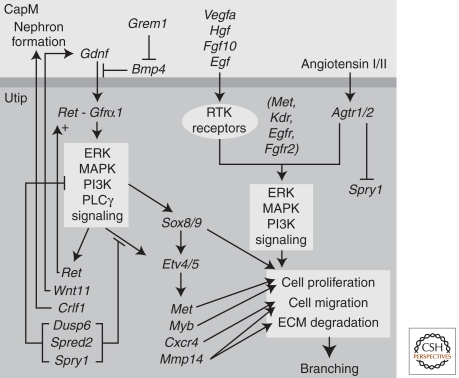
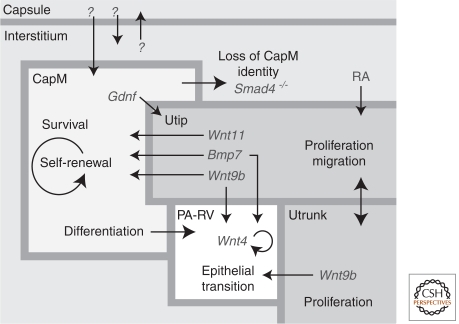
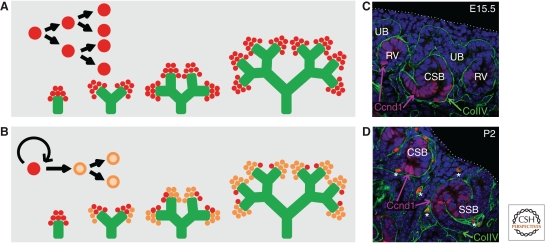
The mammalian kidney is a vital organ with considerable cellular complexity and functional diversity. Kidney development is notable for requiring distinct but coincident tubulogenic processes involving reciprocal inductive signals between mesenchymal and epithelial progenitor compartments. Key molecular pathways mediating these interactions have been identified. Further, advances in the analysis of gene expression and gene activity, coupled with a detailed knowledge of cell origins, are enhancing our understanding of kidney morphogenesis and unraveling the normal processes of postnatal repair and identifying disease-causing mechanisms. This article focuses on recent insights into central regulatory processes governing organ assembly and renal disease, and predicts future directions for the field.
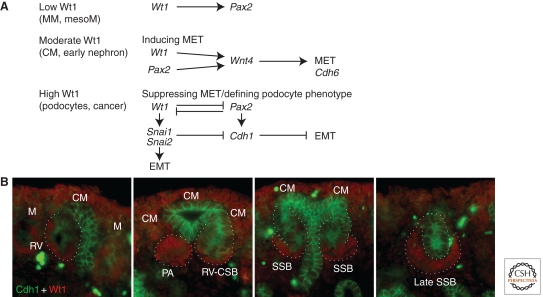
Figures
References
-
- Abrahamson DR, Robert B 2003. Derivation and differentiation of glomerular endothelial cells. Nephrol Dial Transplant 18: vi2–vi7 - PubMed
-
- Angers S, Moon RT 2009. Proximal events in Wnt signal transduction. Nat Rev Mol Cell Biol 10: 468–477 - PubMed
-
- Ballermann BJ 2005. Glomerular endothelial cell differentiation. Kidney Int 67: 1668–1671 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical