

Deregulated telomere transcription causes replication-dependent telomere shortening and promotes cellular senescence
- PMID: 22553368
- PMCID: PMC3413150
- DOI: 10.1093/nar/gks358
Deregulated telomere transcription causes replication-dependent telomere shortening and promotes cellular senescence
Abstract
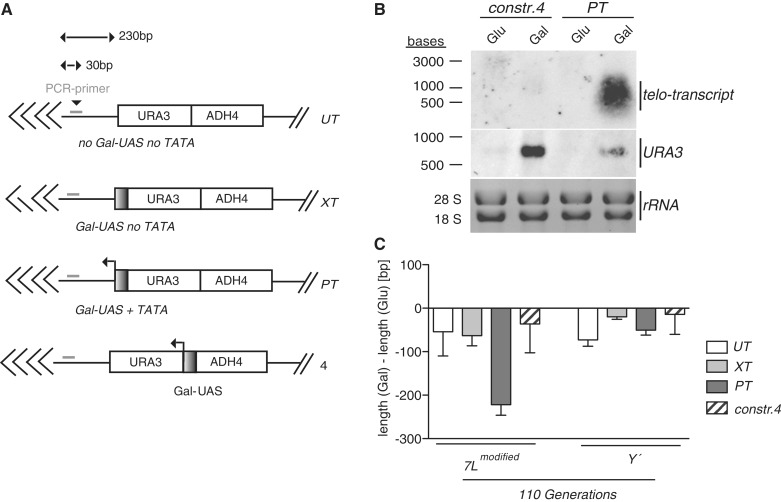
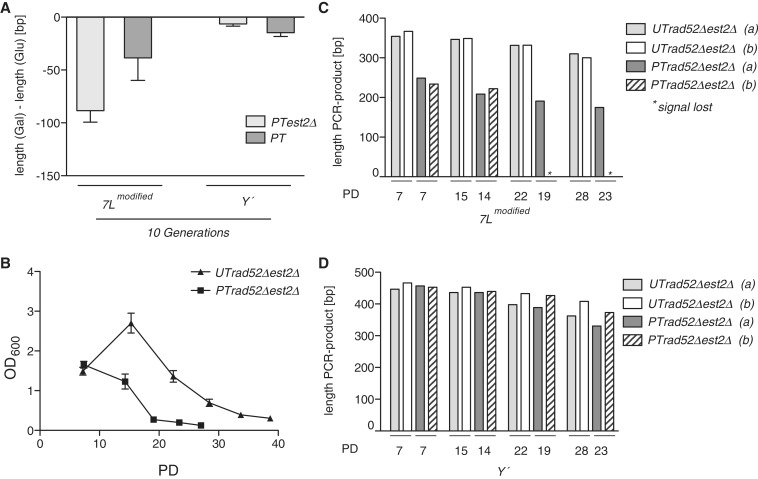
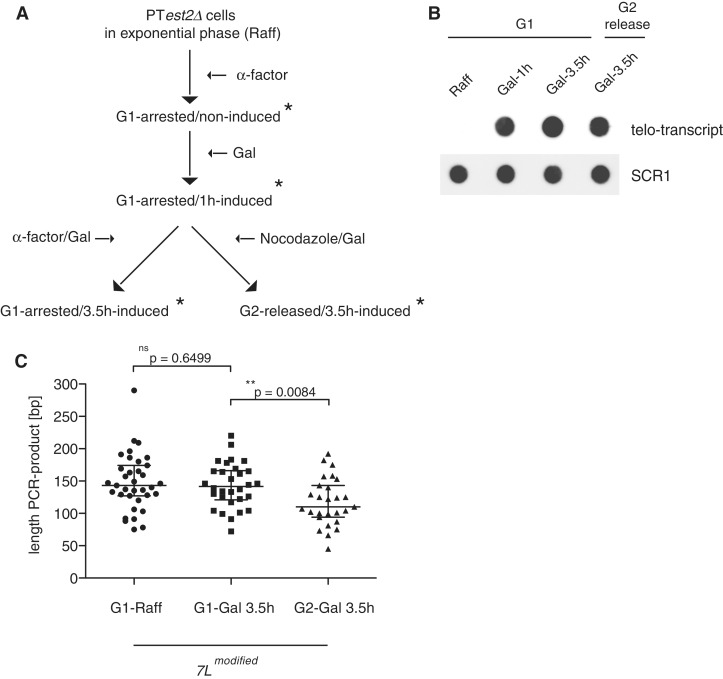
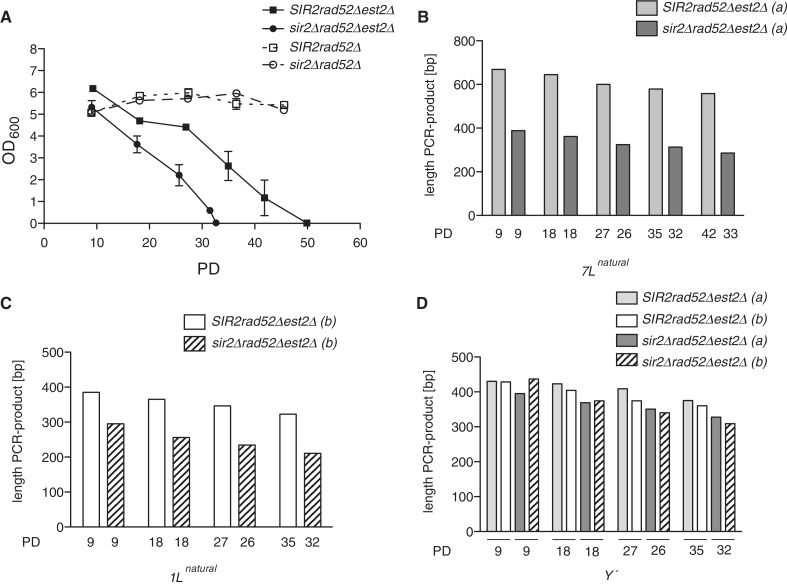
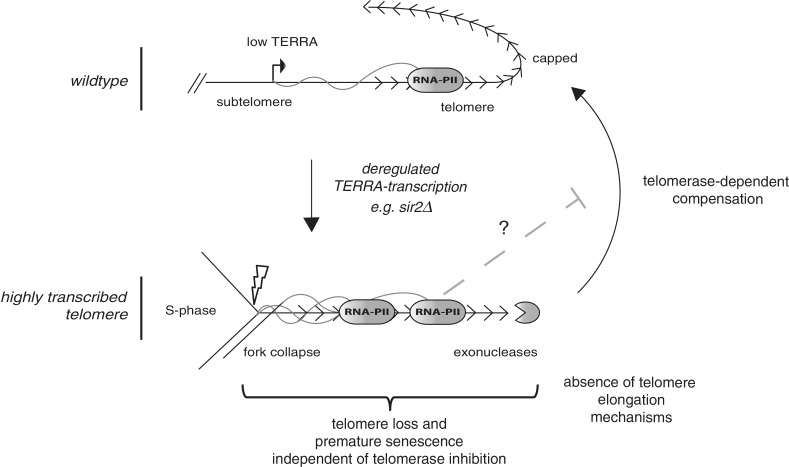
Telomeres are transcribed into non-coding TElomeric Repeat containing RNAs (TERRA). We have employed a transcriptionally inducible telomere to investigate how telomere transcription affects telomere function in Saccharomyces cerevisiae. We report that telomere shortening resulting from high levels of telomere transcription stems from a DNA replication-dependent loss of telomere tracts, which can occur independent of both telomerase inhibition and homologous recombination. We show that in order for telomere loss to occur, transcription must pass through the telomere tract itself producing a TERRA molecule. We demonstrate that increased telomere transcription of a single telomere leads to a premature cellular senescence in the absence of a telomere maintenance mechanism (telomerase and homology directed repair). Similar rapid senescence and telomere shortening are also seen in sir2Δ cells with compromised telomere maintenance, where TERRA levels are increased at natural telomeres. These data suggest that telomere transcription must be tightly controlled to prevent telomere loss and early onset senescence.
Figures
References
-
- Lingner J, Cooper JP, Cech TR. Telomerase and DNA end replication: no longer a lagging strand problem? Science. 1995;269:1533–1534. - PubMed
-
- McEachern MJ, Haber JE. Break-induced replication and recombinational telomere elongation in yeast. Annu. Rev. Biochem. 2006;75:111–135. - PubMed
-
- Lundblad V, Szostak JW. A mutant with a defect in telomere elongation leads to senescence in yeast. Cell. 1989;57:633–643. - PubMed
-
- Palladino F, Laroche T, Gilson E, Axelrod A, Pillus L, Gasser SM. SIR3 and SIR4 proteins are required for the positioning and integrity of yeast telomeres. Cell. 1993;75:543–555. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
