

The putative protein methyltransferase LAE1 controls cellulase gene expression in Trichoderma reesei
- PMID: 22554051
- PMCID: PMC3370264
- DOI: 10.1111/j.1365-2958.2012.08083.x
The putative protein methyltransferase LAE1 controls cellulase gene expression in Trichoderma reesei
Abstract
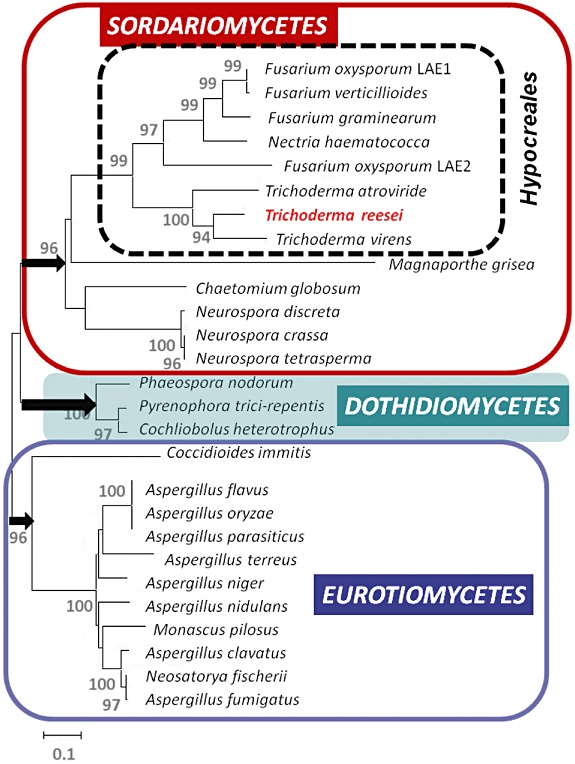
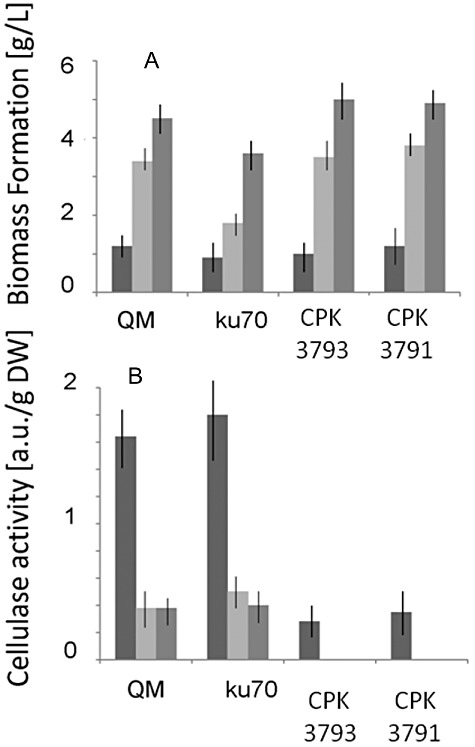
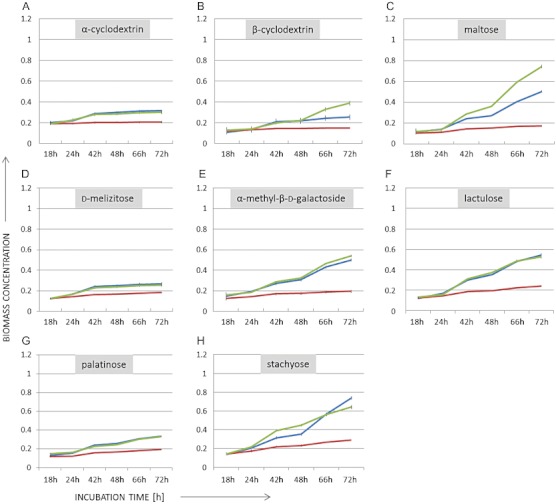
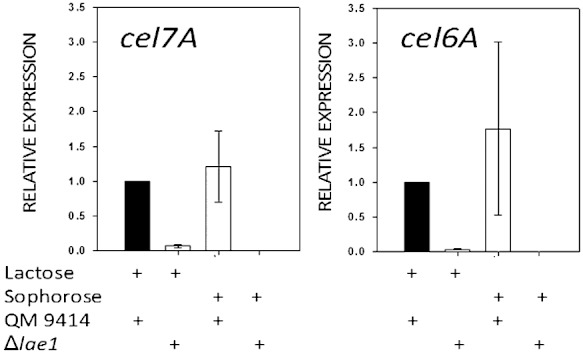
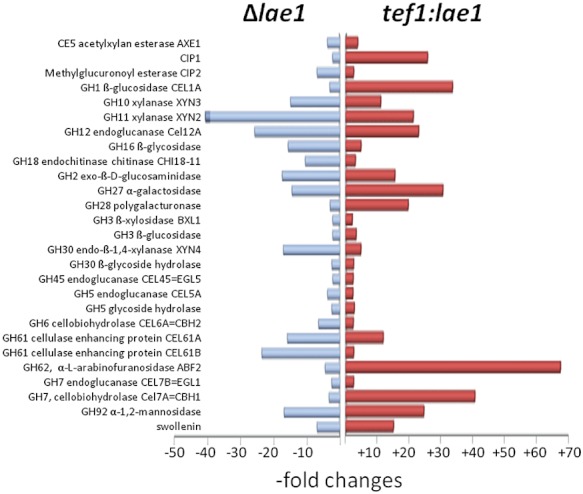
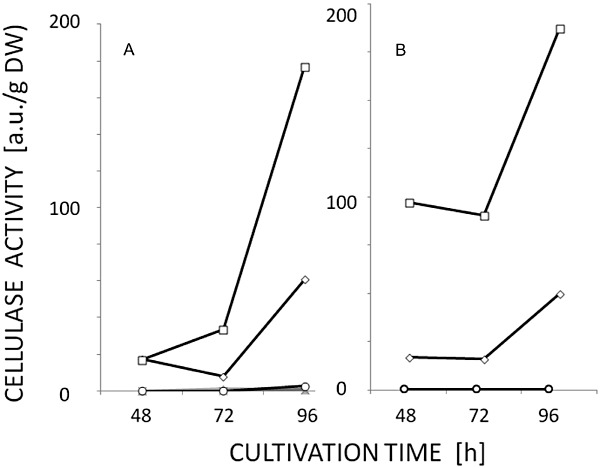
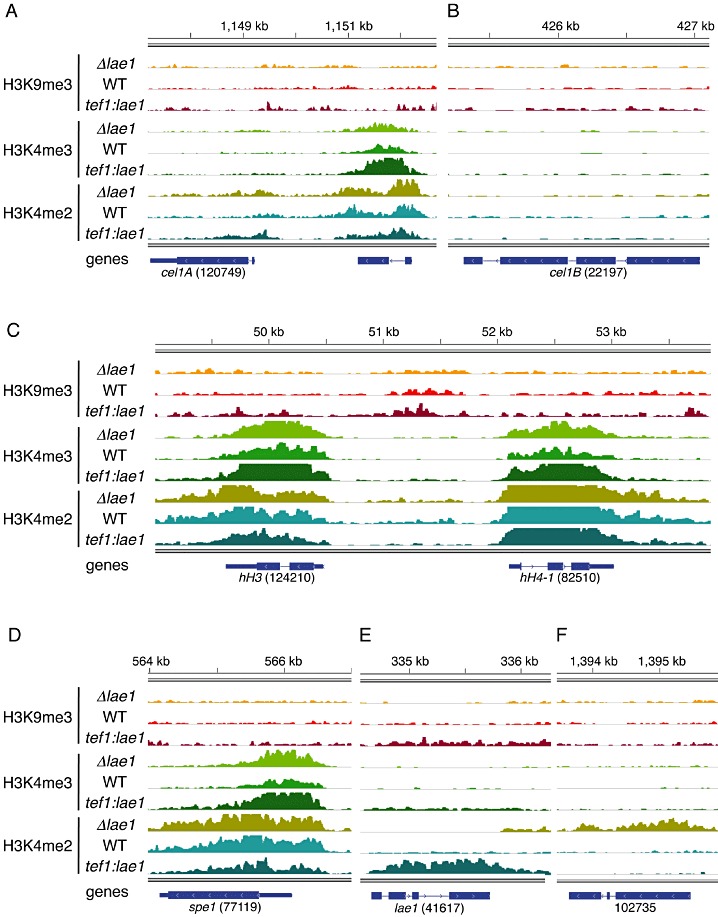
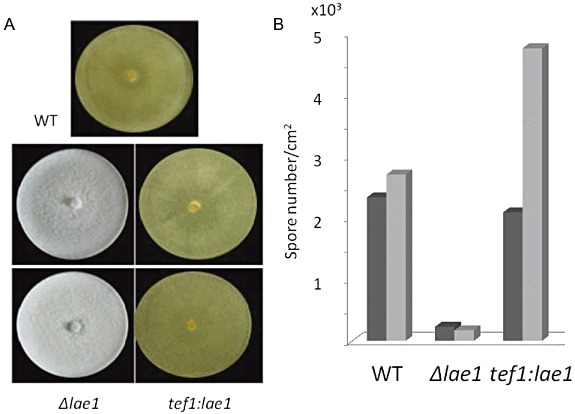
Trichoderma reesei is an industrial producer of enzymes that degrade lignocellulosic polysaccharides to soluble monomers, which can be fermented to biofuels. Here we show that the expression of genes for lignocellulose degradation are controlled by the orthologous T. reesei protein methyltransferase LAE1. In a lae1 deletion mutant we observed a complete loss of expression of all seven cellulases, auxiliary factors for cellulose degradation, β-glucosidases and xylanases were no longer expressed. Conversely, enhanced expression of lae1 resulted in significantly increased cellulase gene transcription. Lae1-modulated cellulase gene expression was dependent on the function of the general cellulase regulator XYR1, but also xyr1 expression was LAE1-dependent. LAE1 was also essential for conidiation of T. reesei. Chromatin immunoprecipitation followed by high-throughput sequencing ('ChIP-seq') showed that lae1 expression was not obviously correlated with H3K4 di- or trimethylation (indicative of active transcription) or H3K9 trimethylation (typical for heterochromatin regions) in CAZyme coding regions, suggesting that LAE1 does not affect CAZyme gene expression by directly modulating H3K4 or H3K9 methylation. Our data demonstrate that the putative protein methyltransferase LAE1 is essential for cellulase gene expression in T. reesei through mechanisms that remain to be identified.
© 2012 Blackwell Publishing Ltd.
Figures
References
-
- Bayram O, Braus GH. Coordination of secondary metabolism and development in fungi: the velvet family of regulatory proteins. FEMS Microbiol Rev. 2012;36:1–24. - PubMed
-
- Bayram O, Krappmann S, Ni M, Bok JW, Helmstaedt K, Valerius O, et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science. 2008;320:1504–1506. - PubMed
-
- Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Ser B. 1995;57:289–300.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
