

DNA aptamers to human immunodeficiency virus reverse transcriptase selected by a primer-free SELEX method: characterization and comparison with other aptamers
- PMID: 22554064
- PMCID: PMC3423876
- DOI: 10.1089/nat.2011.0327
DNA aptamers to human immunodeficiency virus reverse transcriptase selected by a primer-free SELEX method: characterization and comparison with other aptamers
Abstract
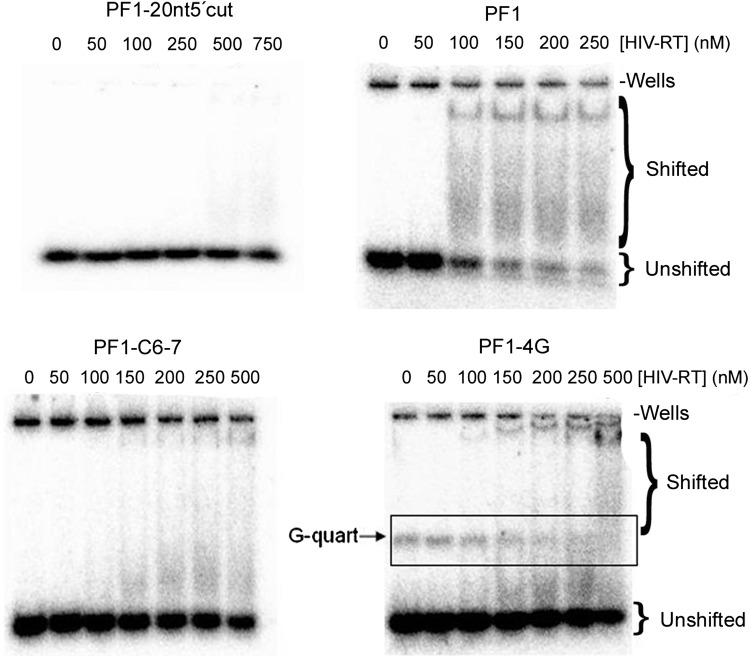
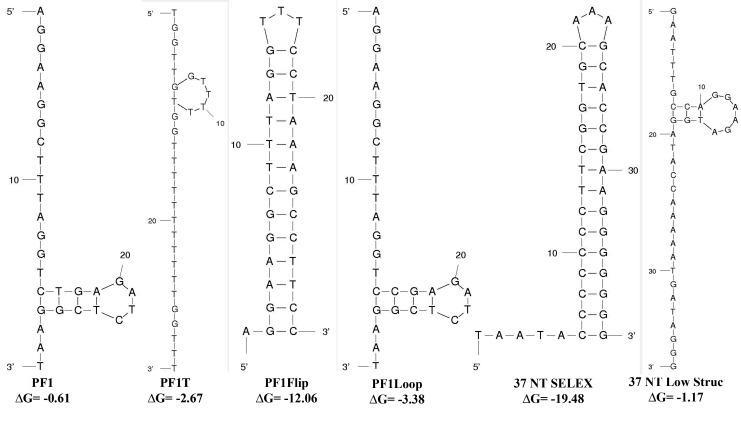
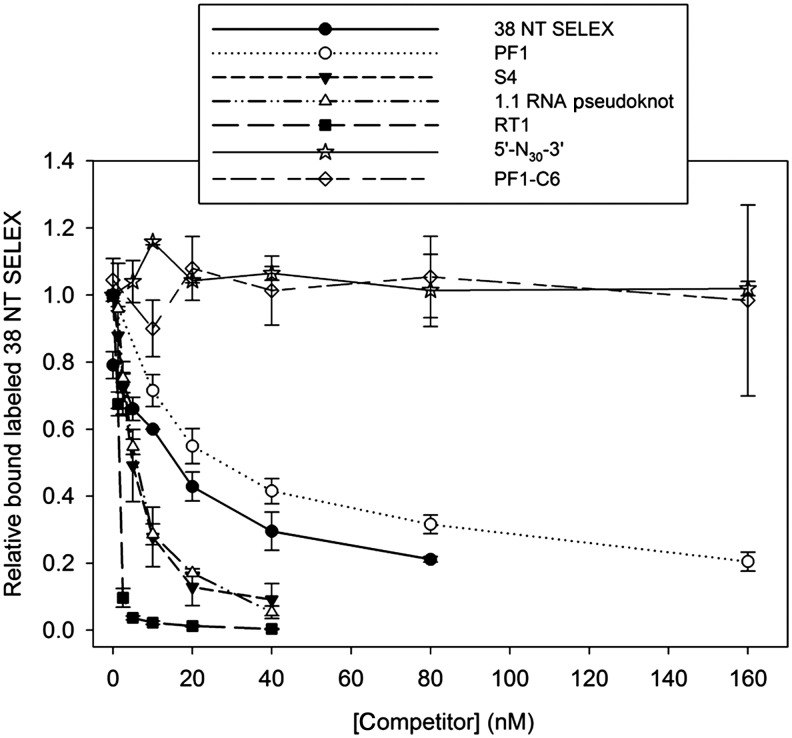
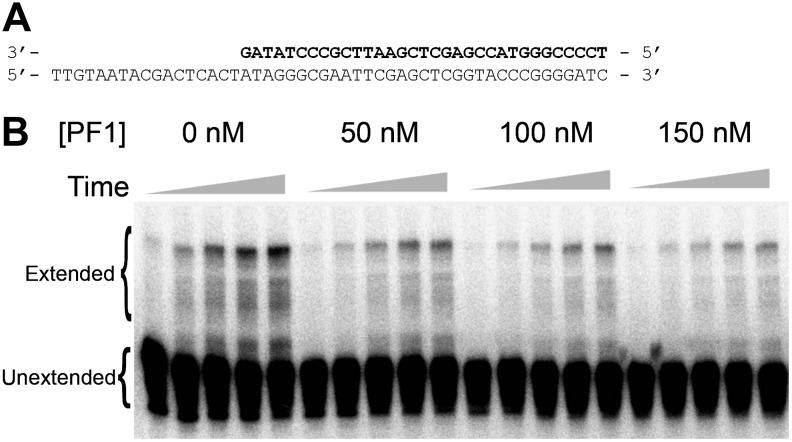
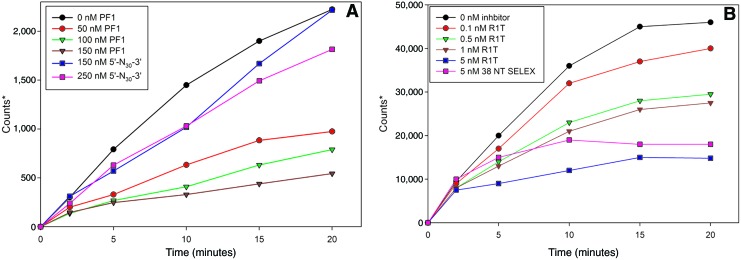
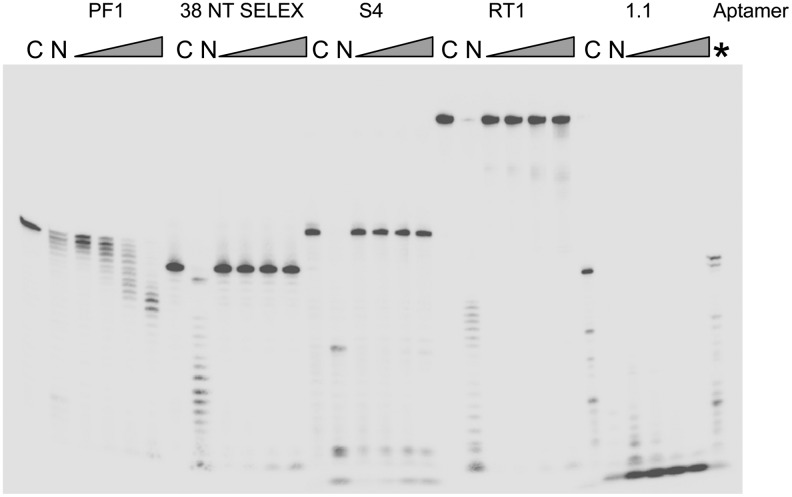
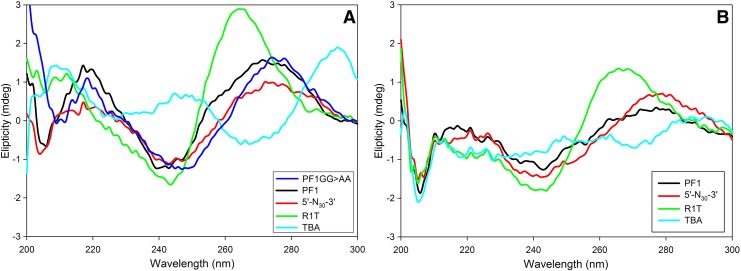
A 30-nucleotide DNA aptamer (5'-AGGAAGGCTTTAGGTCTGAGATCTCGGAAT-3', denoted PF1) selected for high affinity to human immunodeficiency virus reverse transcriptase (HIV RT) using a primer-free SELEX (systematic evolution of ligands by exponential enrichment) method was characterized to determine features promoting tight binding. PF1's equilibrium dissociation constant for RT was ∼80 nM, over 10-fold lower than a random 30-mer. Changing the 2 terminal diguanosine repeats (underlined above) to diadenosine or dithymidine modestly decreased binding. Any changes to the 2 central diguanosines dramatically decreased binding. Binding was highly sensitive to length, with any truncations that deleted part of the 4 diguanosine motifs resulting in a 6-fold or more decrease in affinity. Even a construct with all the diguanosine motifs but lacking the 5' terminal A and 3 nucleotides at the 3' end showed ∼3-fold binding decrease. Changes to the nucleotides between the diguanosines, even those that did not alter PF1's low secondary structure (free energy of folding ΔG=-0.61 kcal/mol), dramatically decreased binding, suggesting sequence specificity. Despite the diguanosine motifs, circular dichroism (CD) spectra indicated that PF1 did not form a G-quartet. PF1 inhibited HIV RT synthesis with a half-maximal inhibitory value (IC(50)) of ∼60 nM. Larger, more structured RT DNA aptamers based on the HIV polypurine tract and those that formed G-quartets (denoted S4 and R1T) were more potent inhibitors, with IC(50) values of ∼4 and ∼1 nM, respectively. An RNA pseudoknot aptamer (denoted 1.1) showed an IC(50) near 4 nM. Competition binding assays with PF1 and several previously characterized RT aptamers indicated that they all bound at or near the primer-template pocket. These other more structured and typically larger aptamers bound more tightly than PF1 to RT based on filter binding assays. Results indicate that PF1 represents a new class of RT aptamers that are relatively small and have very low secondary structure, attributes that could be advantageous for further development as HIV inhibitors.
Figures
References
-
- ANDREOLA M.L. PILEUR F. CALMELS C. VENTURA M. TARRAGO-LITVAK L. TOULME J.J. LITVAK S. DNA aptamers selected against the HIV-1 RNase H display in vitro antiviral activity. Biochemistry. 2001;40:10087–10094. - PubMed
-
- BERNARDI F. GAGGELLI E. MOLTENI E. PORCIATTI E. VALENSIN D. VALENSIN G. 1H and 13C-NMR and molecular dynamics studies of cyclosporin a interacting with magnesium(II) or cerium(III) in acetonitrile. Conformational changes and cis-trans conversion of peptide bonds. Biophys. J. 2006;90:1350–1361. - PMC - PubMed
-
- BOCK L.C. GRIFFIN L.C. LATHAM J.A. VERMAAS E.H. TOOLE J.J. Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature. 1992;355:564–566. - PubMed
-
- BRODY E.N. GOLD L. Aptamers as therapeutic and diagnostic agents. J. Biotechnol. 2000;74:5–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
