

Expanding the genetic code of Caenorhabditis elegans using bacterial aminoacyl-tRNA synthetase/tRNA pairs
- PMID: 22554080
- PMCID: PMC3401359
- DOI: 10.1021/cb200542j
Expanding the genetic code of Caenorhabditis elegans using bacterial aminoacyl-tRNA synthetase/tRNA pairs
Abstract
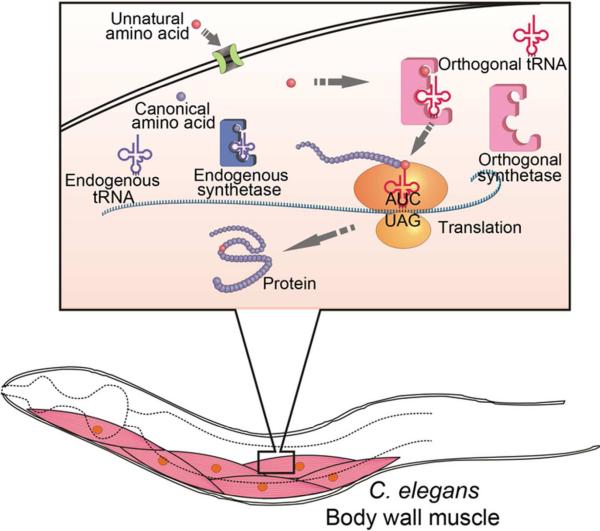
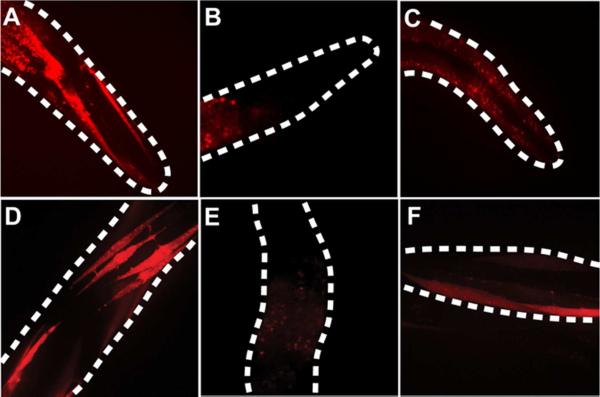
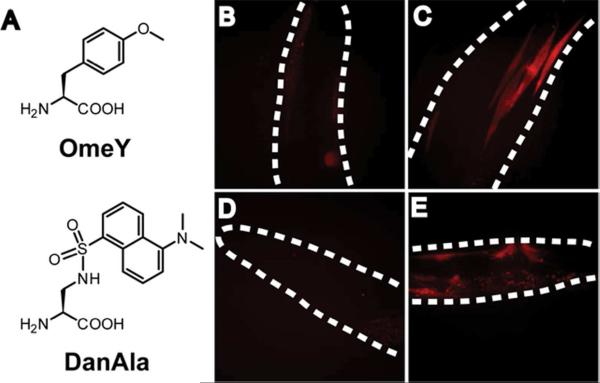
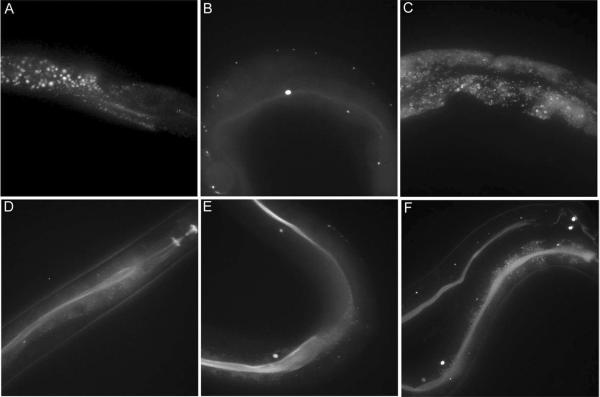
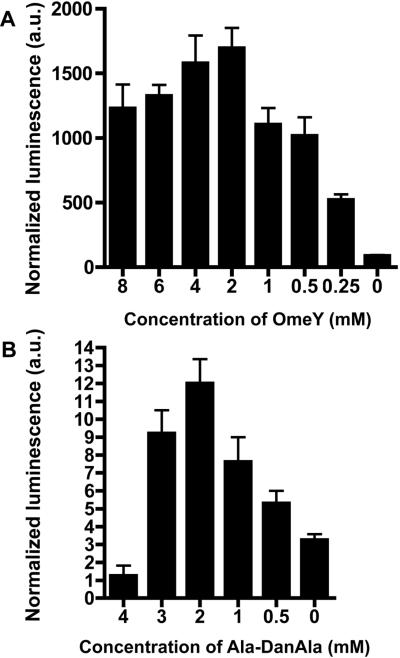
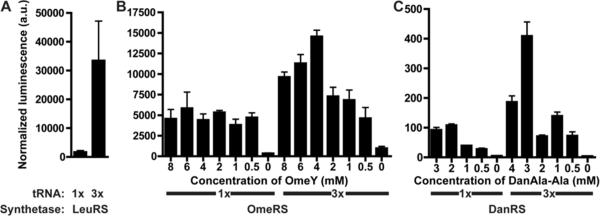
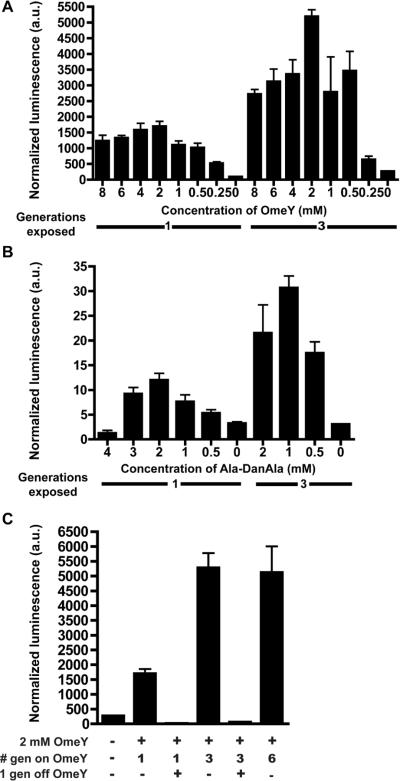
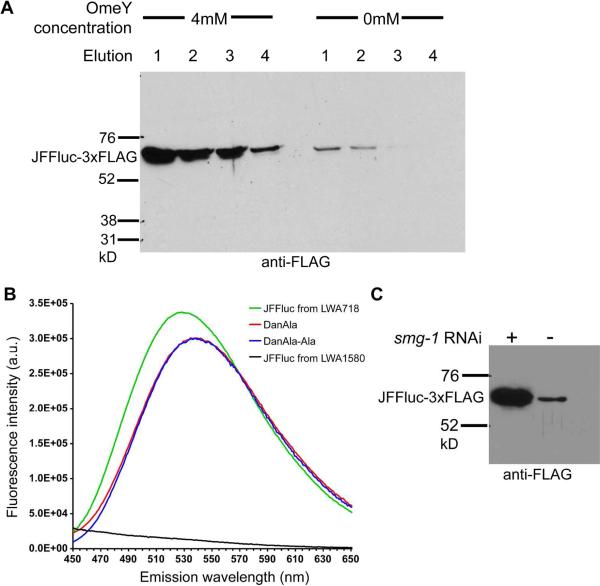
The genetic code specifies 20 common amino acids and is largely preserved in both single and multicellular organisms. Unnatural amino acids (Uaas) have been genetically incorporated into proteins by using engineered orthogonal tRNA/aminoacyl-tRNA synthetase (RS) pairs, enabling new research capabilities and precision inaccessible with common amino acids. We show here that Escherichia coli tyrosyl and leucyl amber suppressor tRNA/RS pairs can be evolved to incorporate different Uaas in response to the amber stop codon UAG into various proteins in Caenorhabditis elegans. To accurately report Uaa incorporation in worms, we found that it is crucial to integrate the UAG-containing reporter gene into the genome rather than to express it on an extrachromosomal array from which variable expression can lead to reporter activation independent of the amber-suppressing tRNA/RS. Synthesizing a Uaa in a dipeptide drives Uaa uptake and bioavailability. Uaa incorporation has dosage, temporal, tRNA copy, and temperature dependencies similar to those of endogenous amber suppression. Uaa incorporation efficiency was improved by impairing the nonsense-mediated mRNA decay pathway through knockdown of smg-1. We have generated stable transgenic worms capable of genetically encoding Uaas, enabling Uaa exploitation to address complex biological problems within a metazoan. We anticipate our strategies will be generally extendable to other multicellular organisms.
Figures
References
-
- Wang L, Brock A, Herberich B, Schultz PG. Expanding the genetic code of Escherichia coli. Science. 2001;292:498–500. - PubMed
-
- Chin JW, Cropp TA, Anderson JC, Mukherji M, Zhang Z, Schultz PG. An expanded eukaryotic genetic code. Science. 2003;301:964–967. - PubMed
-
- Wang Q, Wang L. New methods enabling efficient incorporation of unnatural amino acids in yeast. J. Am. Chem. Soc. 2008;130:6066–6067. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
