

Structure, function, and assembly of heme centers in mitochondrial respiratory complexes
- PMID: 22554985
- PMCID: PMC3601904
- DOI: 10.1016/j.bbamcr.2012.04.008
Structure, function, and assembly of heme centers in mitochondrial respiratory complexes
Abstract
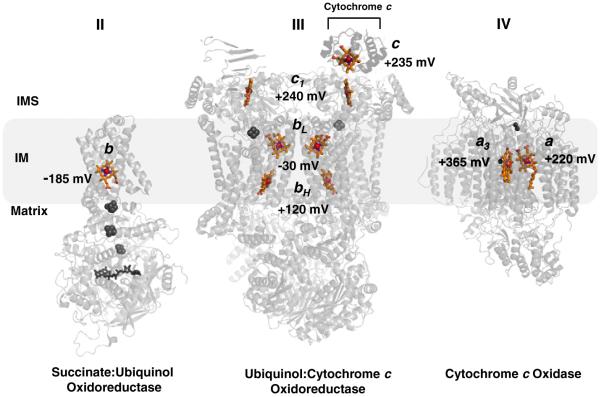
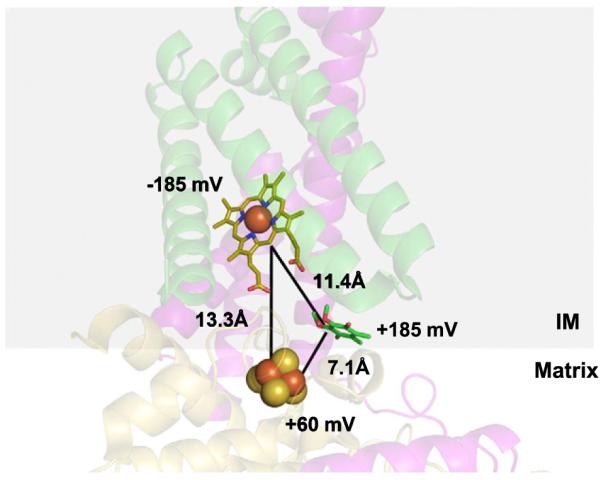
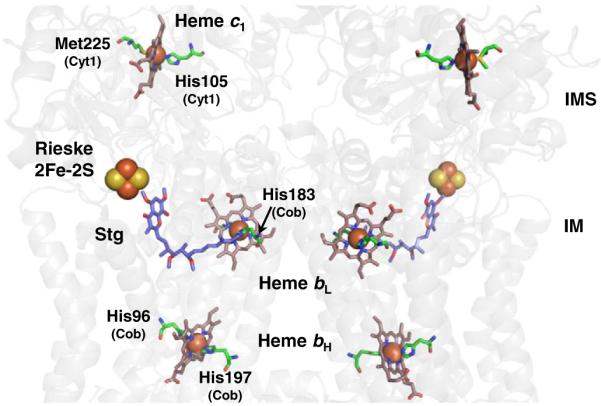
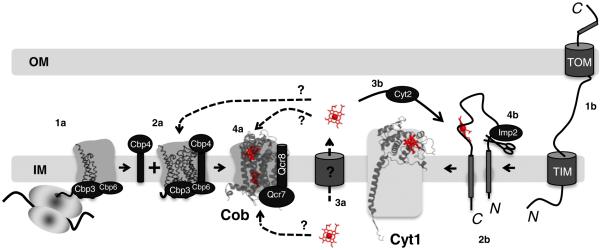
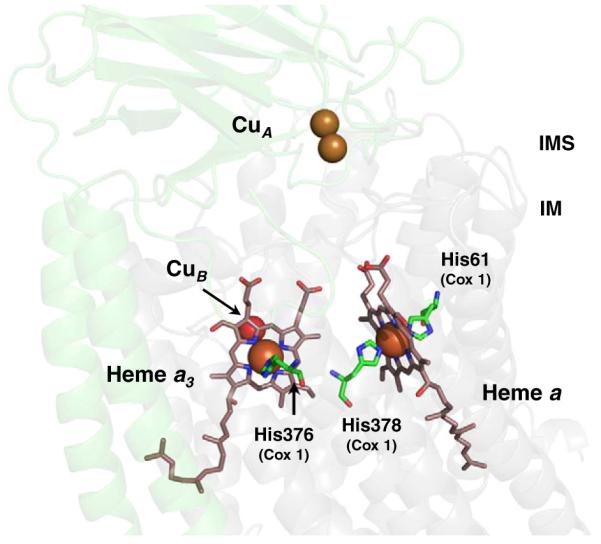
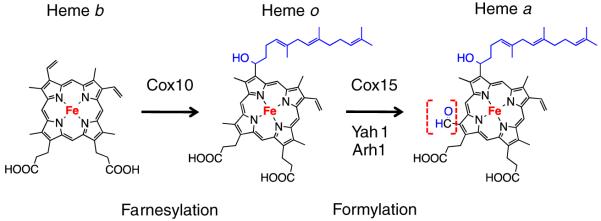
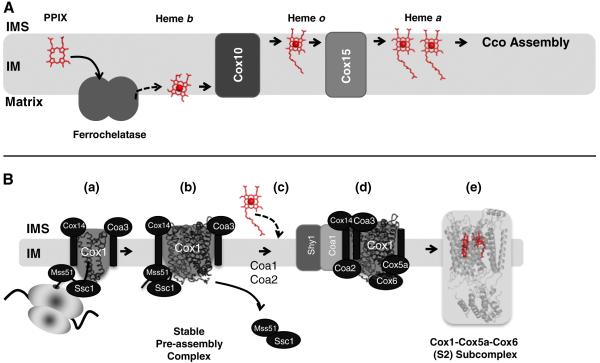
The sequential flow of electrons in the respiratory chain, from a low reduction potential substrate to O(2), is mediated by protein-bound redox cofactors. In mitochondria, hemes-together with flavin, iron-sulfur, and copper cofactors-mediate this multi-electron transfer. Hemes, in three different forms, are used as a protein-bound prosthetic group in succinate dehydrogenase (complex II), in bc(1) complex (complex III) and in cytochrome c oxidase (complex IV). The exact function of heme b in complex II is still unclear, and lags behind in operational detail that is available for the hemes of complex III and IV. The two b hemes of complex III participate in the unique bifurcation of electron flow from the oxidation of ubiquinol, while heme c of the cytochrome c subunit, Cyt1, transfers these electrons to the peripheral cytochrome c. The unique heme a(3), with Cu(B), form a catalytic site in complex IV that binds and reduces molecular oxygen. In addition to providing catalytic and electron transfer operations, hemes also serve a critical role in the assembly of these respiratory complexes, which is just beginning to be understood. In the absence of heme, the assembly of complex II is impaired, especially in mammalian cells. In complex III, a covalent attachment of the heme to apo-Cyt1 is a prerequisite for the complete assembly of bc(1), whereas in complex IV, heme a is required for the proper folding of the Cox 1 subunit and subsequent assembly. In this review, we provide further details of the aforementioned processes with respect to the hemes of the mitochondrial respiratory complexes. This article is part of a Special Issue entitled: Cell Biology of Metals.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Antonini E, Brunori M. Hemoglobin and Myoglobin in their Reactions with Ligands. Elsevier; Amsterdamn and New York: 1971.
-
- Rodgers KR. Heme-based sensors in biological systems. Curr. Opin. Chem. Biol. 1999;3:158–167. - PubMed
-
- Sono M, Roach MP, Coulter ED, Dawson JH. Heme-containing oxygenases. Chem. Rev. 1996;96:2841–2888. - PubMed
-
- Mense SM, Zhang L. Heme: a versatile signaling molecule controlling the activities of diverse regulators ranging from transcription factors to MAP kinases. Cell. Res. 2006;16:681–692. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
