

MORC family ATPases required for heterochromatin condensation and gene silencing
- PMID: 22555433
- PMCID: PMC3376212
- DOI: 10.1126/science.1221472
MORC family ATPases required for heterochromatin condensation and gene silencing
Abstract
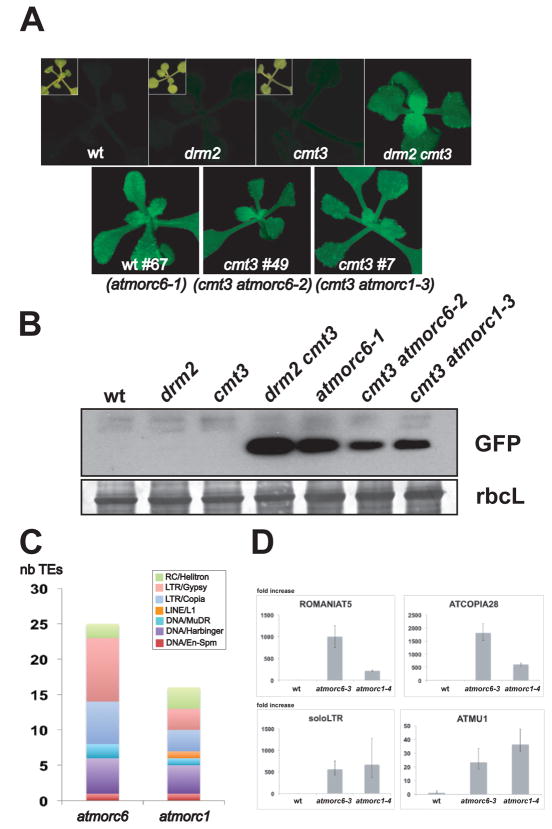
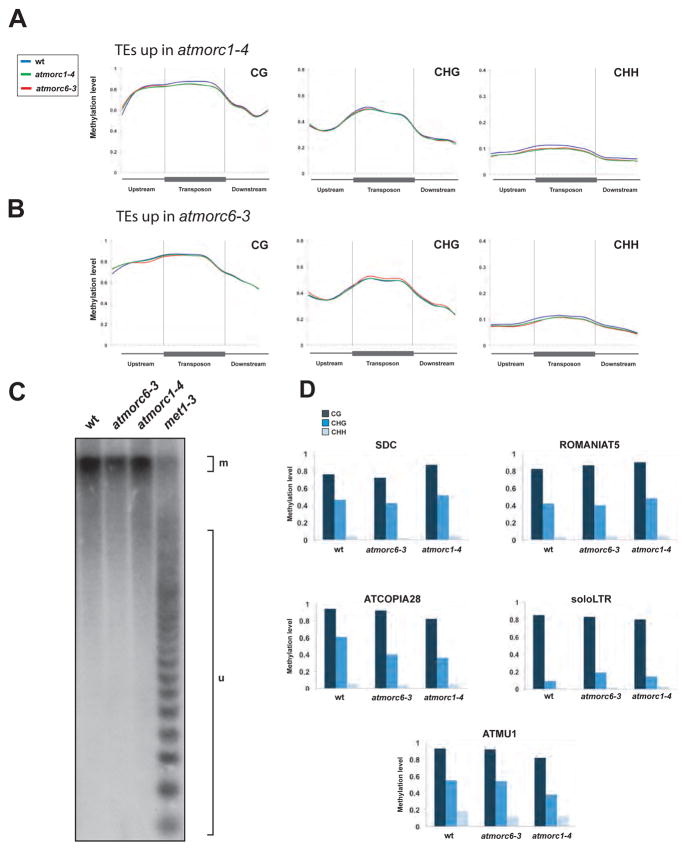
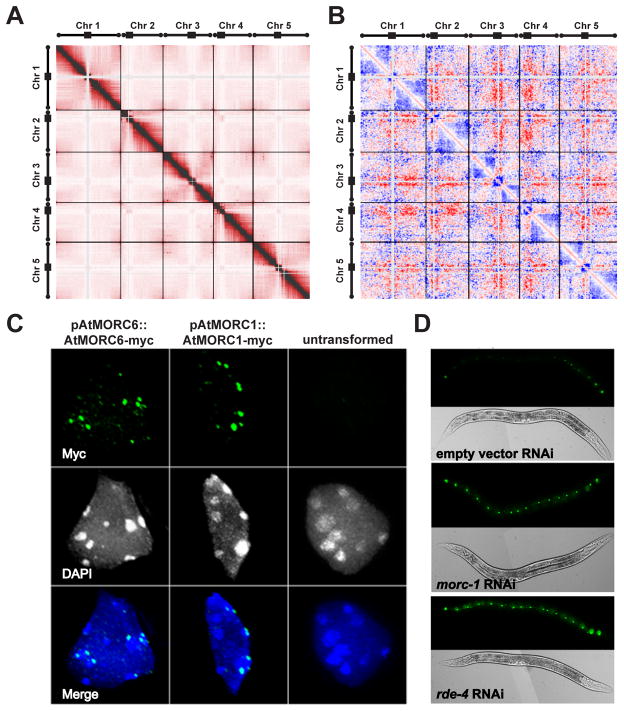
Transposable elements (TEs) and DNA repeats are commonly targeted by DNA and histone methylation to achieve epigenetic gene silencing. We isolated mutations in two Arabidopsis genes, AtMORC1 and AtMORC6, which cause derepression of DNA-methylated genes and TEs but no losses of DNA or histone methylation. AtMORC1 and AtMORC6 are members of the conserved Microrchidia (MORC) adenosine triphosphatase (ATPase) family, which are predicted to catalyze alterations in chromosome superstructure. The atmorc1 and atmorc6 mutants show decondensation of pericentromeric heterochromatin, increased interaction of pericentromeric regions with the rest of the genome, and transcriptional defects that are largely restricted to loci residing in pericentromeric regions. Knockdown of the single MORC homolog in Caenorhabditis elegans also impairs transgene silencing. We propose that the MORC ATPases are conserved regulators of gene silencing in eukaryotes.
Figures
Comment in
-
Molecular biology. All packed up and ready to go.Science. 2012 Jun 15;336(6087):1391-2. doi: 10.1126/science.1224272. Science. 2012. PMID: 22700909 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- HG003143/HG/NHGRI NIH HHS/United States
- GM007185/GM/NIGMS NIH HHS/United States
- R01 HG003143/HG/NHGRI NIH HHS/United States
- GM075060/GM/NIGMS NIH HHS/United States
- T32 GM007544/GM/NIGMS NIH HHS/United States
- R01 GM088565/GM/NIGMS NIH HHS/United States
- F32GM100617/GM/NIGMS NIH HHS/United States
- R01 GM075060/GM/NIGMS NIH HHS/United States
- R37 GM060398/GM/NIGMS NIH HHS/United States
- F32 GM100617/GM/NIGMS NIH HHS/United States
- GM088565/GM/NIGMS NIH HHS/United States
- R01 GM060398/GM/NIGMS NIH HHS/United States
- GM60398/GM/NIGMS NIH HHS/United States
- T32 GM007185/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
