

Structural basis for polyadenosine-RNA binding by Nab2 Zn fingers and its function in mRNA nuclear export
- PMID: 22560733
- PMCID: PMC3384006
- DOI: 10.1016/j.str.2012.03.011
Structural basis for polyadenosine-RNA binding by Nab2 Zn fingers and its function in mRNA nuclear export
Abstract
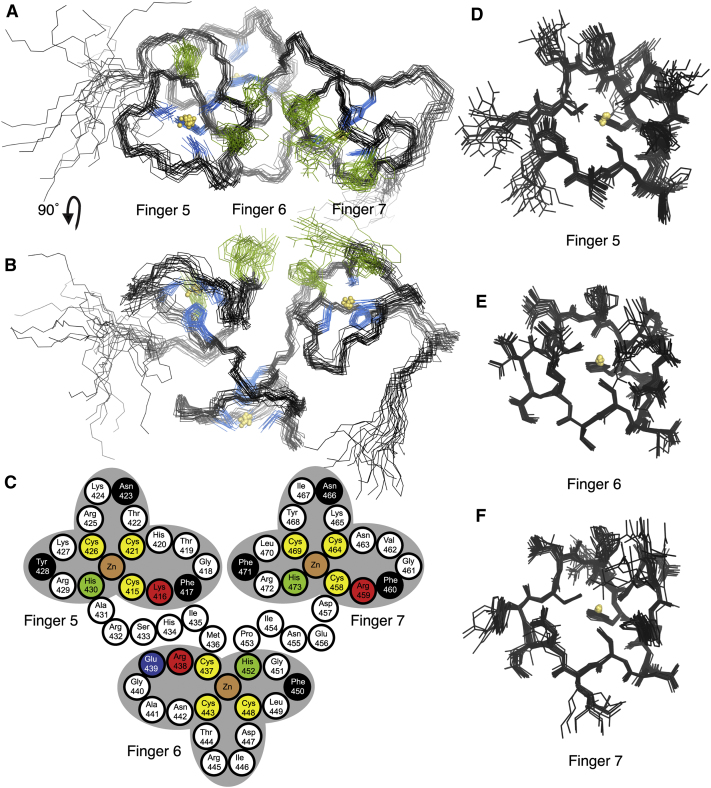
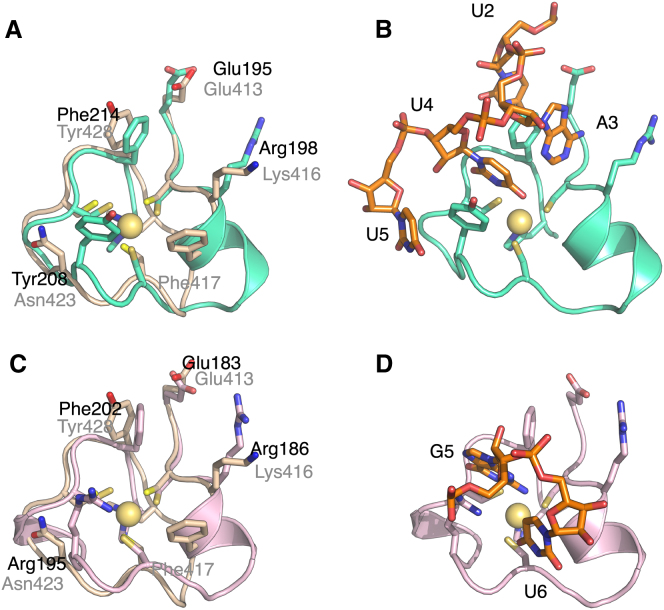
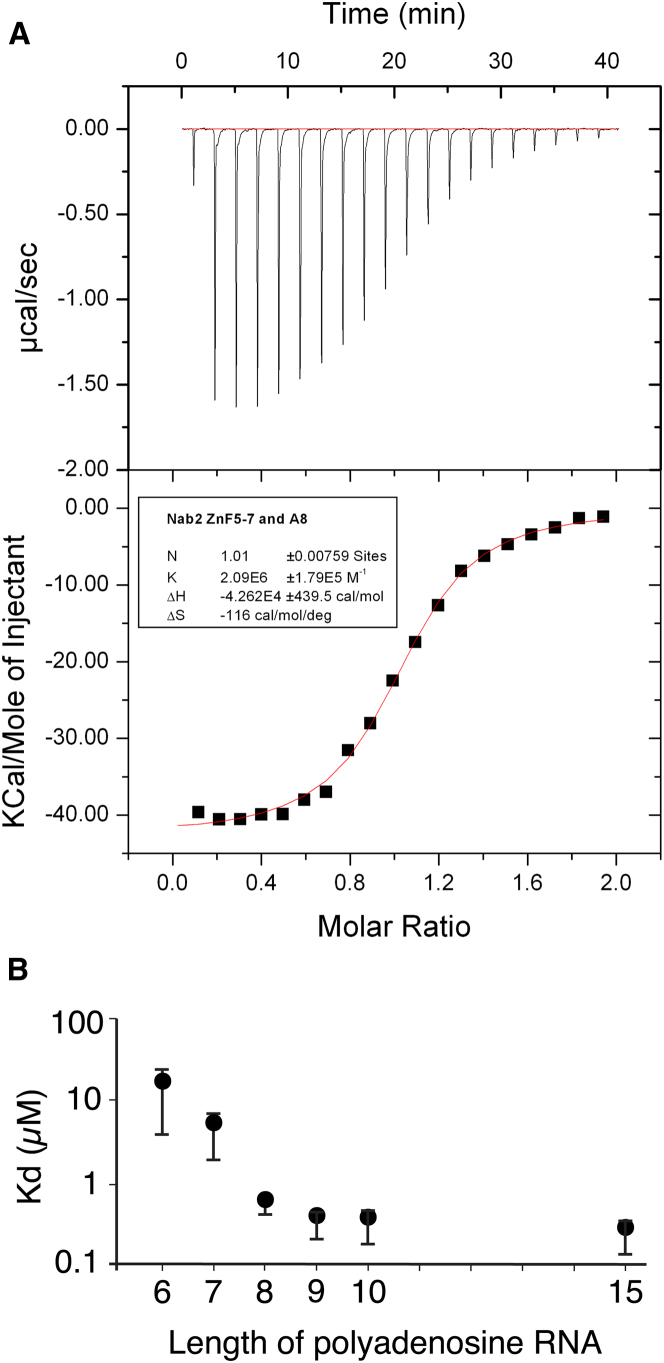
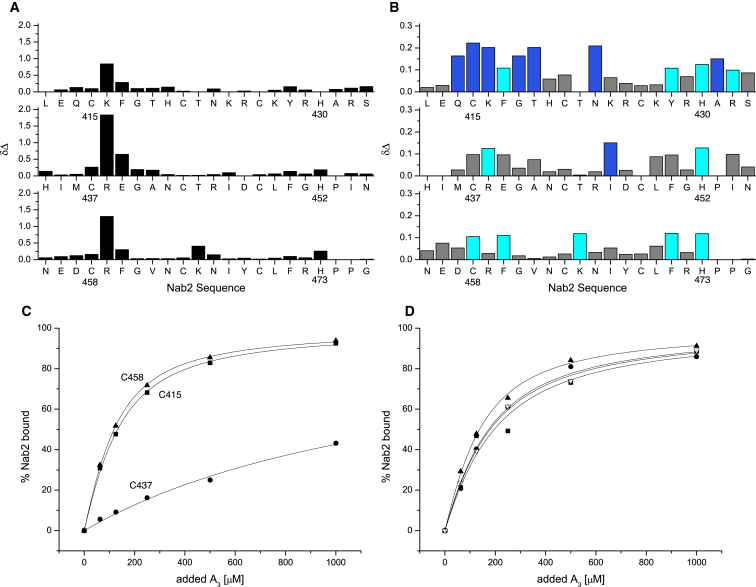
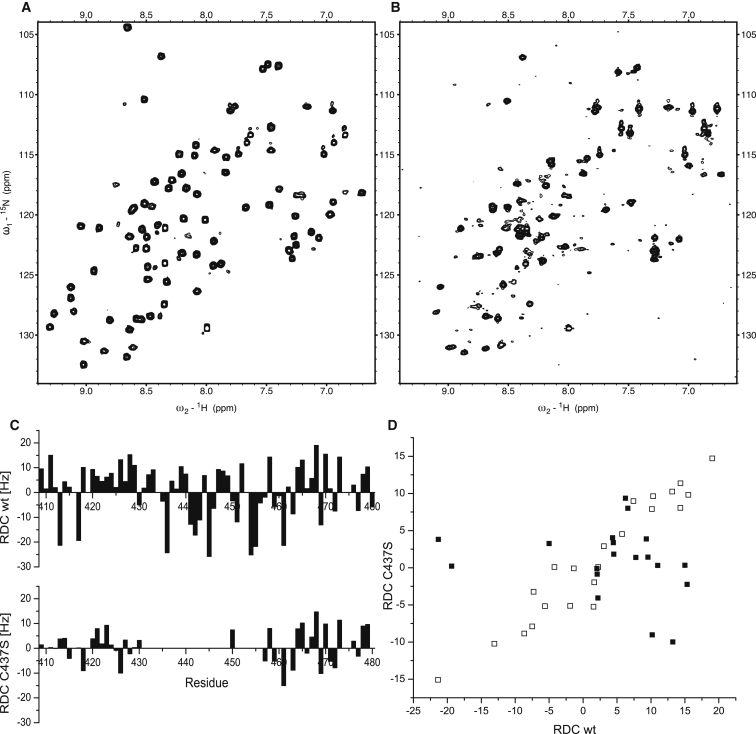
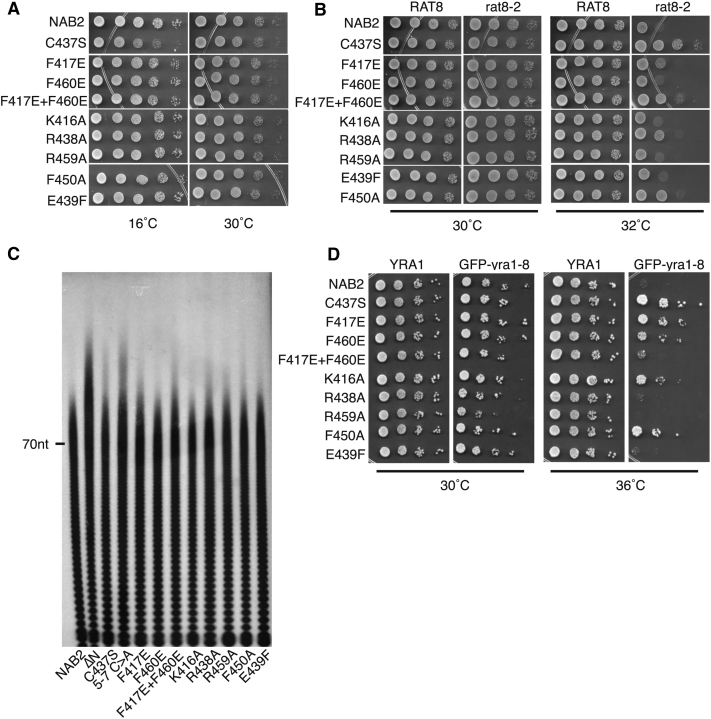
Polyadenylation regulation and efficient nuclear export of mature mRNPs both require the polyadenosine-RNA-binding protein, Nab2, which contains seven CCCH Zn fingers. We describe here the solution structure of fingers 5-7, which are necessary and sufficient for high-affinity polyadenosine-RNA binding, and identify key residues involved. These Zn fingers form a single structural unit. Structural coherence is lost in the RNA-binding compromised Nab2-C437S mutant, which also suppresses the rat8-2 allele of RNA helicase Dbp5. Structure-guided Nab2 variants indicate that dbp5(rat8-2) suppression is more closely linked to hyperadenylation and suppression of mutant alleles of the nuclear RNA export adaptor, Yra1, than to affinity for polyadenosine-RNA. These results indicate that, in addition to modulating polyA tail length, Nab2 has an unanticipated function associated with generating export-competent mRNPs, and that changes within fingers 5-7 lead to suboptimal assembly of mRNP export complexes that are more easily disassembled by Dbp5 upon reaching the cytoplasm.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures

Comment in
-
Nab2, a three-cornered hat to bind a poly(A) tail.Structure. 2012 Jun 6;20(6):937-8. doi: 10.1016/j.str.2012.05.005. Structure. 2012. PMID: 22681898
References
-
- Adams A., Gottschling D.E., Kaiser C.A., Stearns T. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 1997. Methods in Yeast Genetics.
-
- Aitchison J.D., Blobel G., Rout M.P. Kap104p: a karyopherin involved in the nuclear transport of messenger RNA binding proteins. Science. 1996;274:624–627. - PubMed
-
- Alcázar-Román A.R., Tran E.J., Guo S., Wente S.R. Inositol hexakisphosphate and Gle1 activate the DEAD-box protein Dbp5 for nuclear mRNA export. Nat. Cell Biol. 2006;8:711–716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
