

The structure of monoacylglycerol lipase from Bacillus sp. H257 reveals unexpected conservation of the cap architecture between bacterial and human enzymes
- PMID: 22561231
- PMCID: PMC3790968
- DOI: 10.1016/j.bbalip.2012.04.006
The structure of monoacylglycerol lipase from Bacillus sp. H257 reveals unexpected conservation of the cap architecture between bacterial and human enzymes
Abstract
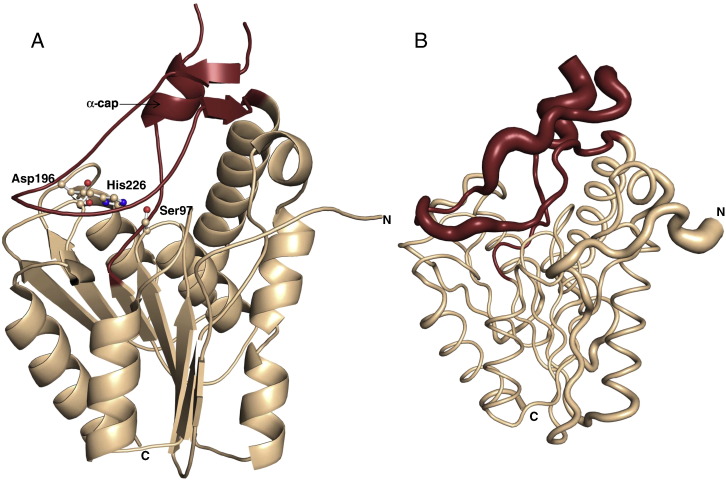
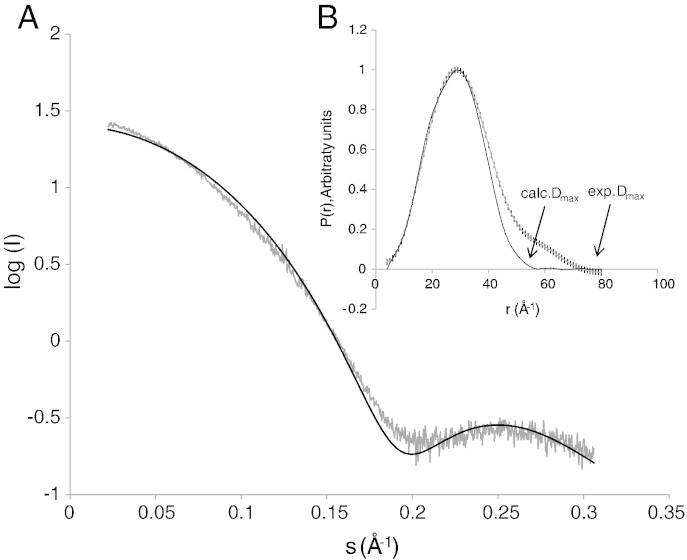
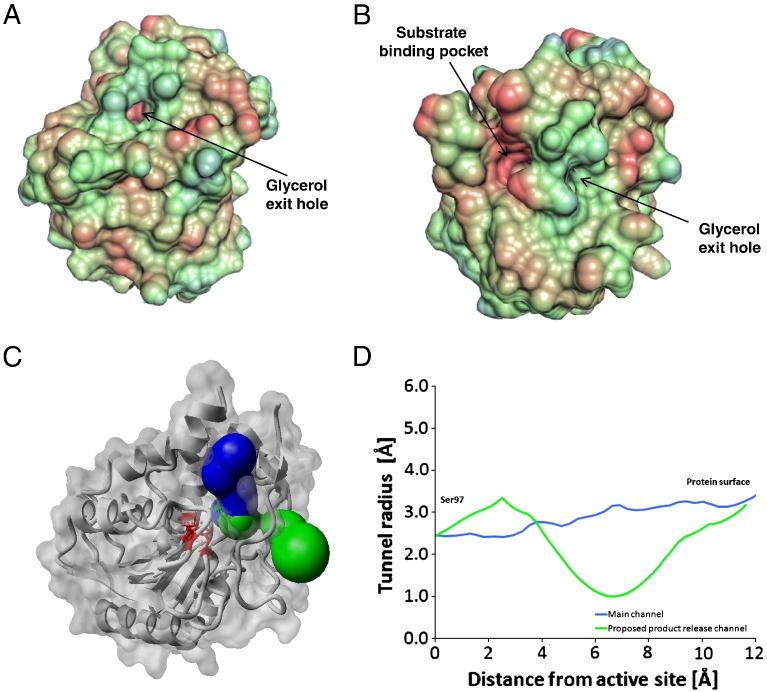
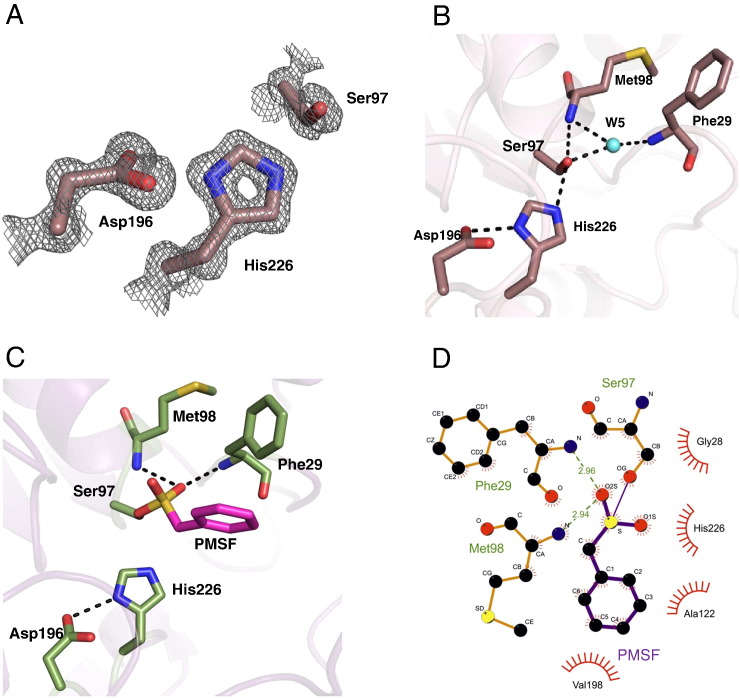
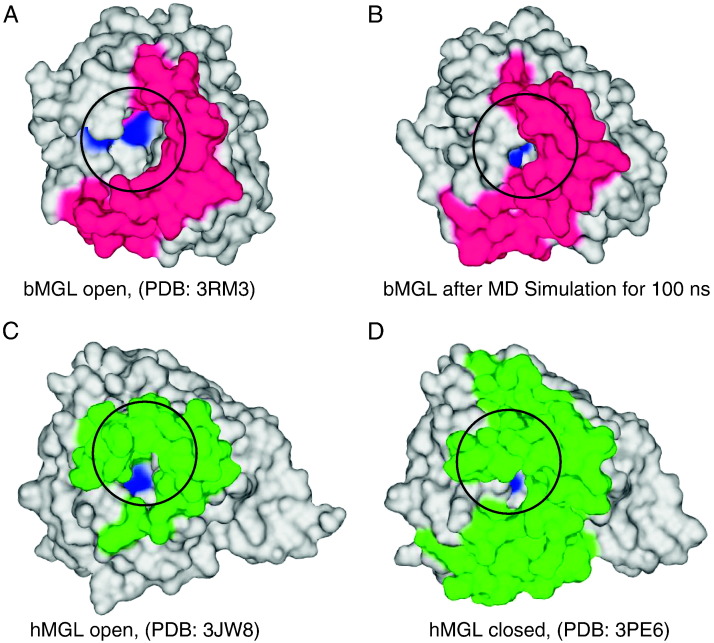
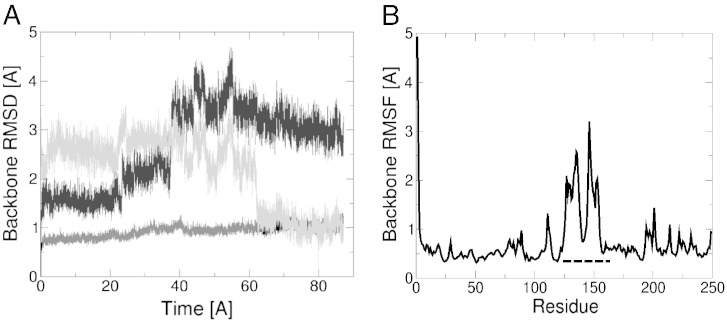
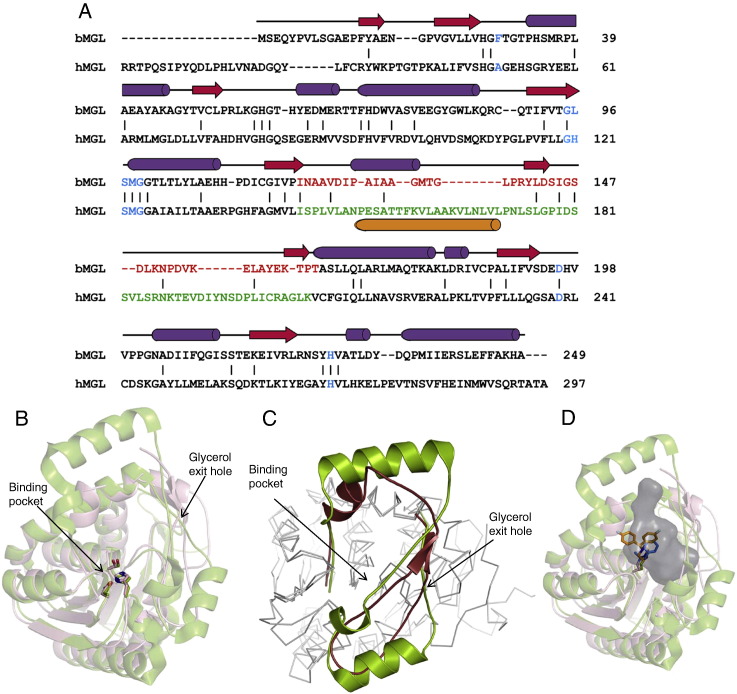
Monoacylglycerol lipases (MGLs) catalyse the hydrolysis of monoacylglycerol into free fatty acid and glycerol. MGLs have been identified throughout all genera of life and have adopted different substrate specificities depending on their physiological role. In humans, MGL plays an integral part in lipid metabolism affecting energy homeostasis, signalling processes and cancer cell progression. In bacteria, MGLs degrade short-chain monoacylglycerols which are otherwise toxic to the organism. We report the crystal structures of MGL from the bacterium Bacillus sp. H257 (bMGL) in its free form at 1.2Å and in complex with phenylmethylsulfonyl fluoride at 1.8Å resolution. In both structures, bMGL adopts an α/β hydrolase fold with a cap in an open conformation. Access to the active site residues, which were unambiguously identified from the protein structure, is facilitated by two different channels. The larger channel constitutes the highly hydrophobic substrate binding pocket with enough room to accommodate monoacylglycerol. The other channel is rather small and resembles the proposed glycerol exit hole in human MGL. Molecular dynamics simulation of bMGL yielded open and closed states of the entrance channel and the glycerol exit hole. Despite differences in the number of residues, secondary structure elements, and low sequence identity in the cap region, this first structure of a bacterial MGL reveals striking structural conservation of the overall cap architecture in comparison with human MGL. Thus it provides insight into the structural conservation of the cap amongst MGLs throughout evolution and provides a framework for rationalising substrate specificities in each organism.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Tornqvist H., Belfrage P. Purification and some properties of a monoacylglycerol-hydrolyzing enzyme of rat adipose tissue. J. Biol. Chem. 1976;251:813–819. - PubMed
-
- Fredrikson G., Tornqvist H., Belfrage P. Hormone-sensitive lipase and monoacylglycerol lipase are both required for complete degradation of adipocyte triacylglycerol. Biochim. Biophys. Acta. 1986;876:288–293. - PubMed
-
- Dinh T.P., Kathuria S., Piomelli D. RNA interference suggests a primary role for monoacylglycerol lipase in the degradation of the endocannabinoid 2-arachidonoylglycerol. Mol. Pharmacol. 2004;66:1260–1264. - PubMed
-
- Di Marzo V., Goparaju S.K., Wang L., Liu J., Batkai S., Jarai Z., Fezza F., Miura G.I., Palmiter R.D., Sugiura T., Kunos G. Leptin-regulated endocannabinoids are involved in maintaining food intake. Nature. 2001;410:822–825. - PubMed
-
- Hohmann A.G., Suplita R.L., Bolton N.M., Neely M.H., Fegley D., Mangieri R., Krey J.F., Walker J.M., Holmes P.V., Crystal J.D., Duranti A., Tontini A., Mor M., Tarzia G., Piomelli D. An endocannabinoid mechanism for stress-induced analgesia. Nature. 2005;435:1108–1112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
