

Cnn1 inhibits the interactions between the KMN complexes of the yeast kinetochore
- PMID: 22561345
- PMCID: PMC3438452
- DOI: 10.1038/ncb2495
Cnn1 inhibits the interactions between the KMN complexes of the yeast kinetochore
Erratum in
- Nat Cell Biol. 2013 Mar;15(3):335
Abstract
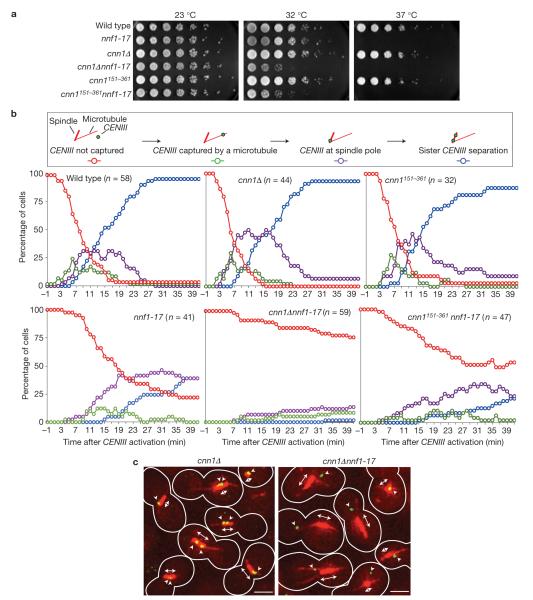
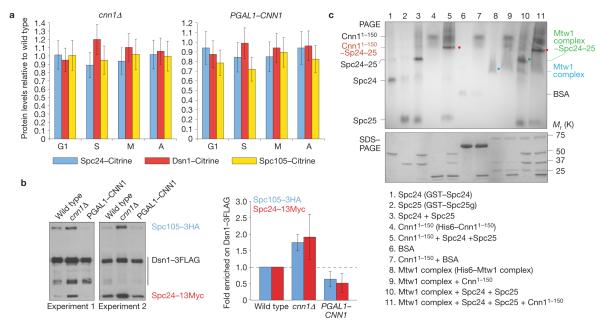
Kinetochores attach the replicated chromosomes to the mitotic spindle and orchestrate their transmission to the daughter cells. Kinetochore-spindle binding and chromosome segregation are mediated by the multi-copy KNL1(Spc105), MIS12(Mtw1) and NDC80(Ndc80) complexes that form the so-called KMN network. KMN-spindle attachment is regulated by the Aurora B(Ipl1) and MPS1(Mps1) kinases. It is unclear whether other mechanisms exist that support KMN activity during the cell cycle. Using budding yeast, we show that kinetochore protein Cnn1 localizes to the base of the Ndc80 complex and promotes a functionally competent configuration of the KMN network. Cnn1 regulates KMN activity in a spatiotemporal manner by inhibiting the interaction between its complexes. Cnn1 activity peaks in anaphase and is driven by the Cdc28, Mps1 and Ipl1 kinases.
Figures
Comment in
-
T time for point centromeres.Nat Cell Biol. 2012 May 30;14(6):559-61. doi: 10.1038/ncb2509. Nat Cell Biol. 2012. PMID: 22561349
References
-
- Lampert F, Westermann S. A blueprint for kinetochores—new insights into the molecular mechanics of cell division. Nat. Rev. Mol. Cell Biol. 2011;12:407–412. - PubMed
-
- Tanaka K, et al. Molecular mechanisms of kinetochore capture by spindle microtubules. Nature. 2005;434:987–994. - PubMed
-
- Cheeseman IM, Chappie JS, Wilson-Kubalek EM, Desai A. The conserved KMN network constitutes the core microtubule-binding site of the kinetochore. Cell. 2006;127:983–997. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
