

Wnt signaling induces proliferation of sensory precursors in the postnatal mouse cochlea
- PMID: 22562792
- PMCID: PMC3361451
- DOI: 10.1073/pnas.1202774109
Wnt signaling induces proliferation of sensory precursors in the postnatal mouse cochlea
Abstract
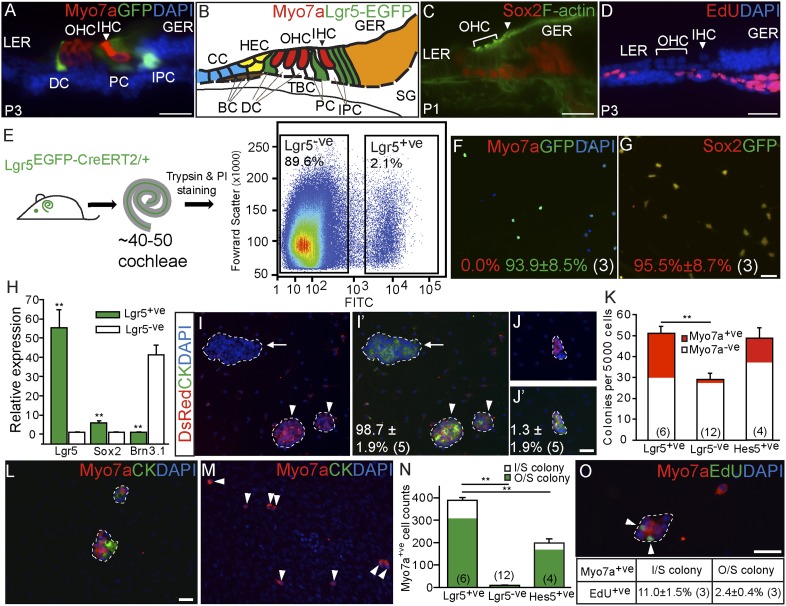
Inner ear hair cells are specialized sensory cells essential for auditory function. Previous studies have shown that the sensory epithelium is postmitotic, but it harbors cells that can behave as progenitor cells in vitro, including the ability to form new hair cells. Lgr5, a Wnt target gene, marks distinct supporting cell types in the neonatal cochlea. Here, we tested the hypothesis that Lgr5(+) cells are Wnt-responsive sensory precursor cells. In contrast to their quiescent in vivo behavior, Lgr5(+) cells isolated by flow cytometry from neonatal Lgr5(EGFP-CreERT2/+) mice proliferated and formed clonal colonies. After 10 d in culture, new sensory cells formed and displayed specific hair cell markers (myo7a, calretinin, parvalbumin, myo6) and stereocilia-like structures expressing F-actin and espin. In comparison with other supporting cells, Lgr5(+) cells were enriched precursors to myo7a(+) cells, most of which formed without mitotic division. Treatment with Wnt agonists increased proliferation and colony-formation capacity. Conversely, small-molecule inhibitors of Wnt signaling suppressed proliferation without compromising the myo7a(+) cells formed by direct differentiation. In vivo lineage tracing supported the idea that Lgr5(+) cells give rise to myo7a(+) hair cells in the neonatal Lgr5(EGFP-CreERT2/+) cochlea. In addition, overexpression of β-catenin initiated proliferation and led to transient expansion of Lgr5(+) cells within the cochlear sensory epithelium. These results suggest that Lgr5 marks sensory precursors and that Wnt signaling can promote their proliferation and provide mechanistic insights into Wnt-responsive progenitor cells during sensory organ development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Cruz RM, Lambert PR, Rubel EW. Light microscopic evidence of hair cell regeneration after gentamicin toxicity in chick cochlea. Arch Otolaryngol Head Neck Surg. 1987;113:1058–1062. - PubMed
-
- Corwin JT, Cotanche DA. Regeneration of sensory hair cells after acoustic trauma. Science. 1988;240:1772–1774. - PubMed
-
- Cafaro J, Lee GS, Stone JS. Atoh1 expression defines activated progenitors and differentiating hair cells during avian hair cell regeneration. Dev Dyn. 2007;236:156–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
