

Differential roles of proteasome and immunoproteasome regulators Pa28αβ, Pa28γ and Pa200 in the degradation of oxidized proteins
- PMID: 22564544
- PMCID: PMC3384713
- DOI: 10.1016/j.abb.2012.04.018
Differential roles of proteasome and immunoproteasome regulators Pa28αβ, Pa28γ and Pa200 in the degradation of oxidized proteins
Abstract
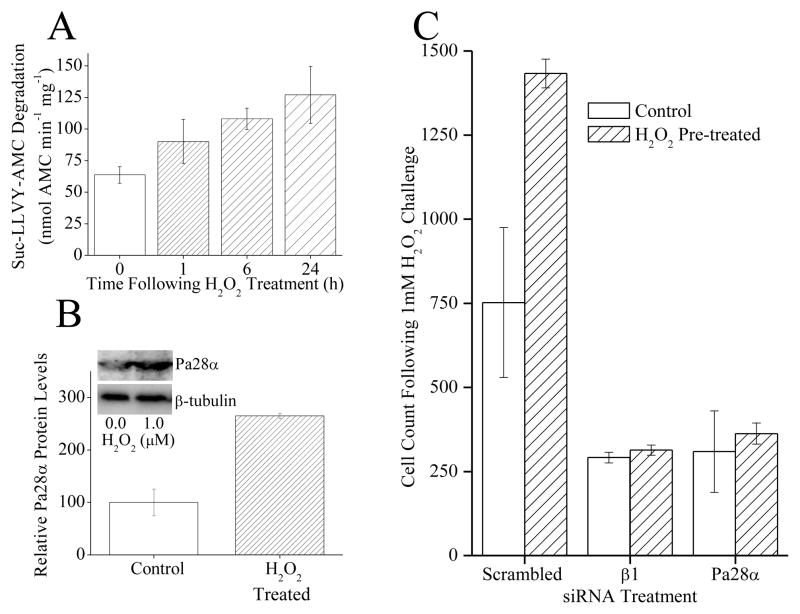
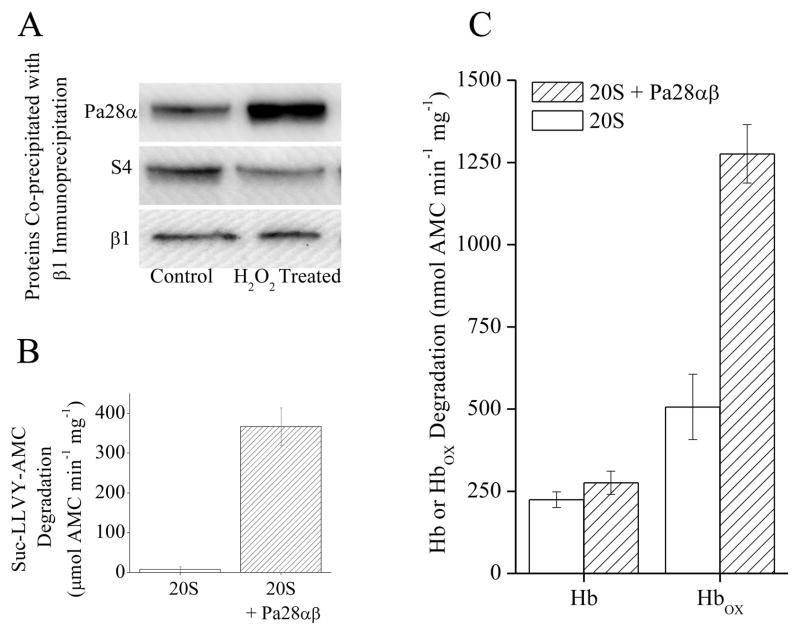
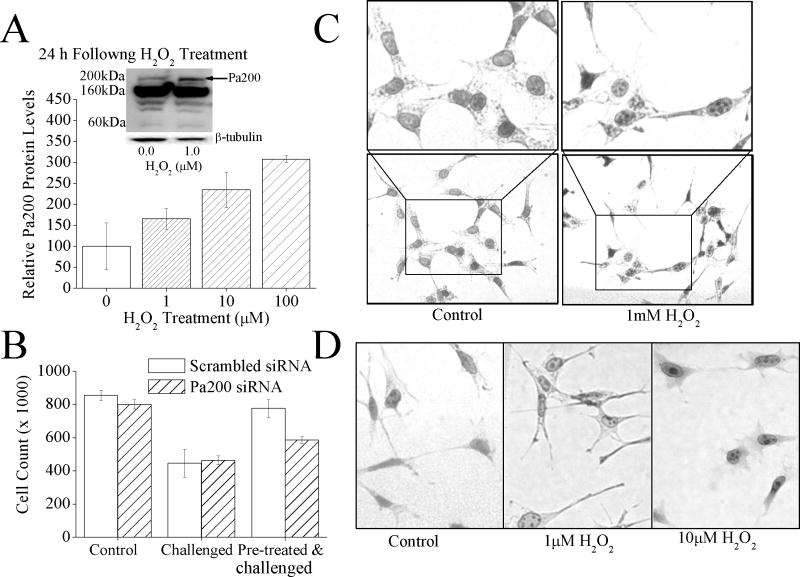
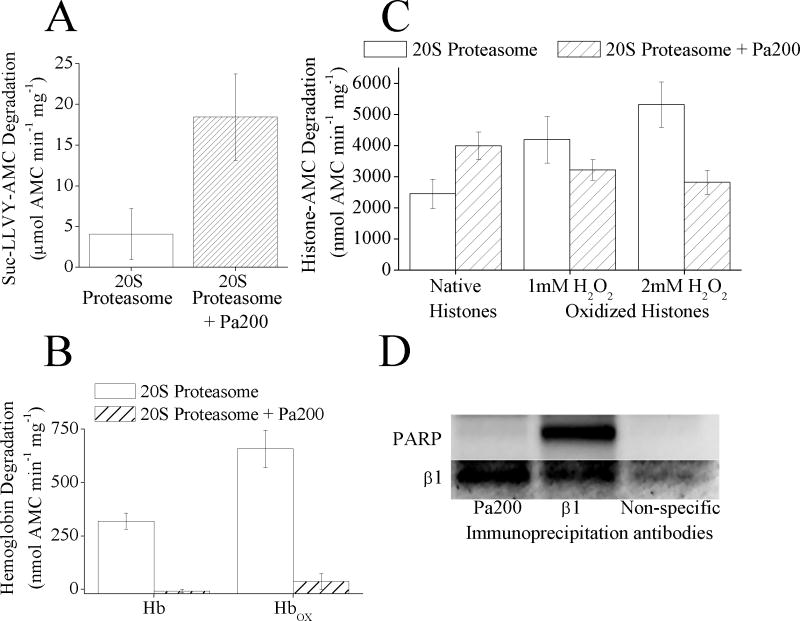
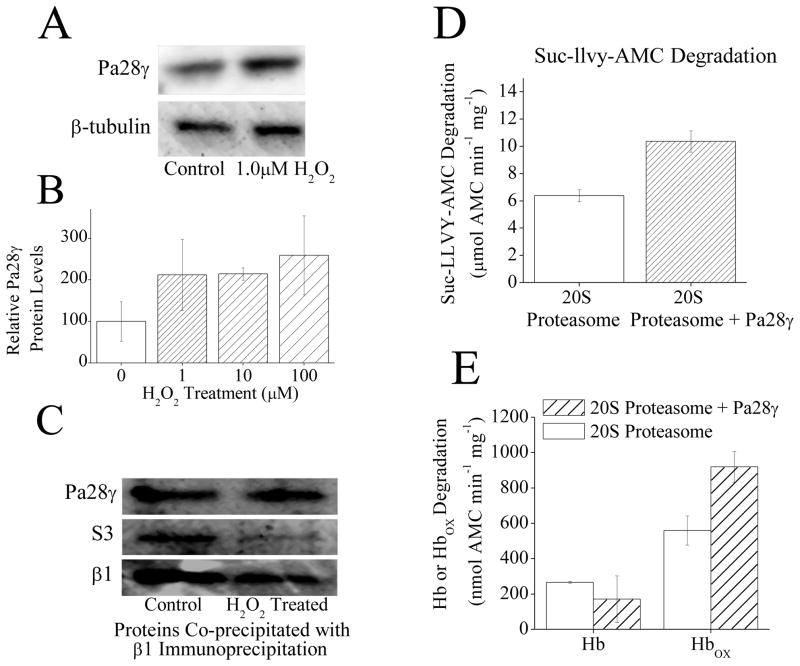
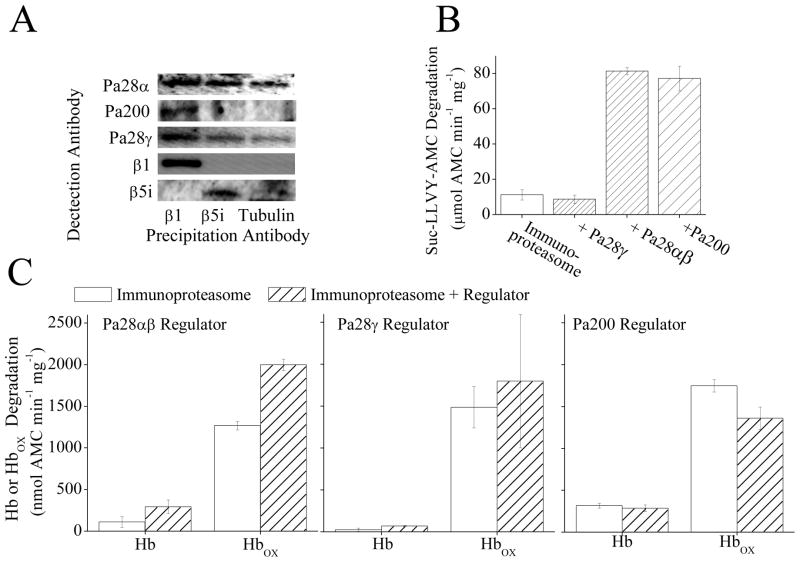
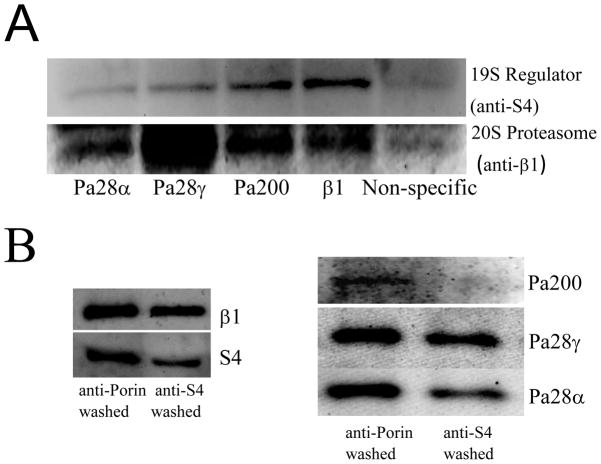
The response and functions of proteasome regulators Pa28αβ (or 11S), Pa28γ and Pa200 in oxidative-stress adaptation (also called hormesis) was studied in murine embryonic fibroblasts (MEFs), using a well-characterized model of cellular adaptation to low concentrations (1.0-10.0 μM) of hydrogen peroxide (H(2)O(2)), which alter gene expression profiles, increasing resistance to higher levels of oxidative-stress. Pa28αβ bound to 20S proteasomes immediately upon H(2)O(2)-treatment, whereas 26S proteasomes were disassembled at the same time. Over the next 24h, the levels of Pa28αβ, Pa28γ and Pa200 proteasome regulators increased during H(2)O(2)-adaptation, whereas the 19S regulator was unchanged. Purified Pa28αβ, and to a lesser extent Pa28γ, significantly increased the ability of purified 20S proteasome to selectively degrade oxidized proteins; Pa28αβ also increased the capacity of purified immunoproteasome to selectively degrade oxidized proteins but Pa28γ did not. Pa200 regulator actually decreased 20S proteasome and immunoproteasome's ability to degrade oxidized proteins but Pa200 and poly-ADP ribose polymerase may cooperate in enabling initiation of DNA repair. Our results indicate that cytoplasmic Pa28αβ and nuclear Pa28γ may both be important regulators of proteasome's ability to degrade oxidatively-damaged proteins, and induced-expression of both 20S proteasome and immunoproteasome, and their Pa28αβ and Pa28γ regulators are important for oxidative-stress adaptation.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
