

Review
doi: 10.1038/embor.2012.55.
Losing balance: the origin and impact of aneuploidy in cancer
Affiliations
- PMID: 22565320
- PMCID: PMC3367240
- DOI: 10.1038/embor.2012.55
Item in Clipboard
Review
Losing balance: the origin and impact of aneuploidy in cancer
EMBO Rep.
.
Abstract
Most solid human tumours are aneuploid, that is, they contain an abnormal number of chromosomes. Paradoxically, however, aneuploidy has been reported to induce a stress response that suppresses cellular proliferation in vitro. Here, we review the progress in our understanding of the causes and effects of aneuploidy in cancer and discuss how, in specific contexts, aneuploidy can provide a growth advantage and facilitate cellular transformation. We also explore the emerging possibilities for targeting the cause or consequences of aneuploidy therapeutically.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
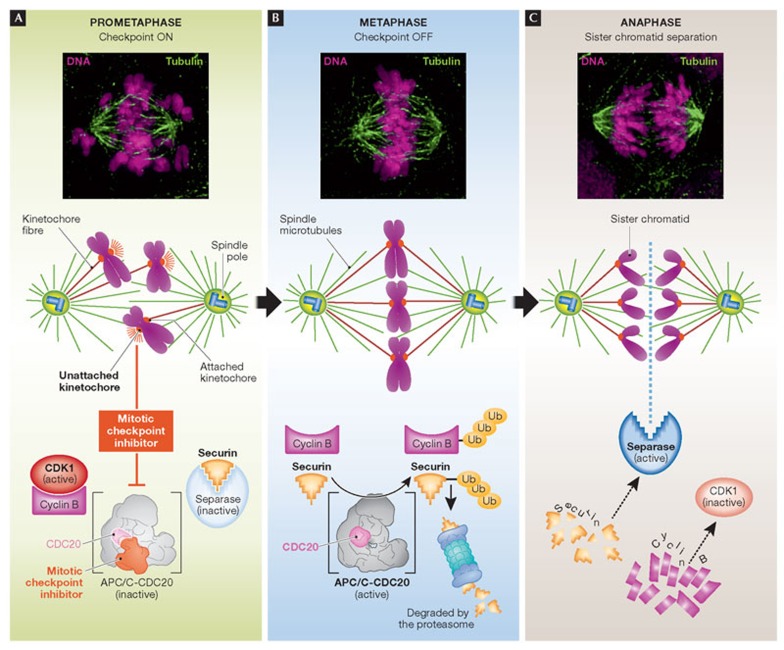
(A) To guard against chromosome missegregation and aneuploidy, cells have evolved a surveillance pathway known as the mitotic checkpoint that halts progression into anaphase until all of the kinetochores have attached to the microtubules of the mitotic spindle. Unattached kinetochores release a diffusible signal that inhibits ubiquitination of cyclin B and securin by the APC/C bound to its activator CDC20. (B) At metaphase, when all kinetochores are correctly attached to microtubules of the spindle, the mitotic checkpoint is silenced and APC/CCDC20 ubiquitinates securin and cyclin B1, thereby targeting them for destruction by the 26S proteasome. (C) Destruction of securin liberates separase, which promotes loss of sister chromatid cohesion, and cyclin B1 destruction inactivates CDK1 thereby promoting mitotic exit. APC/C, anaphase promoting complex/cyclosome; CDK1, cyclin-dependent kinase 1.
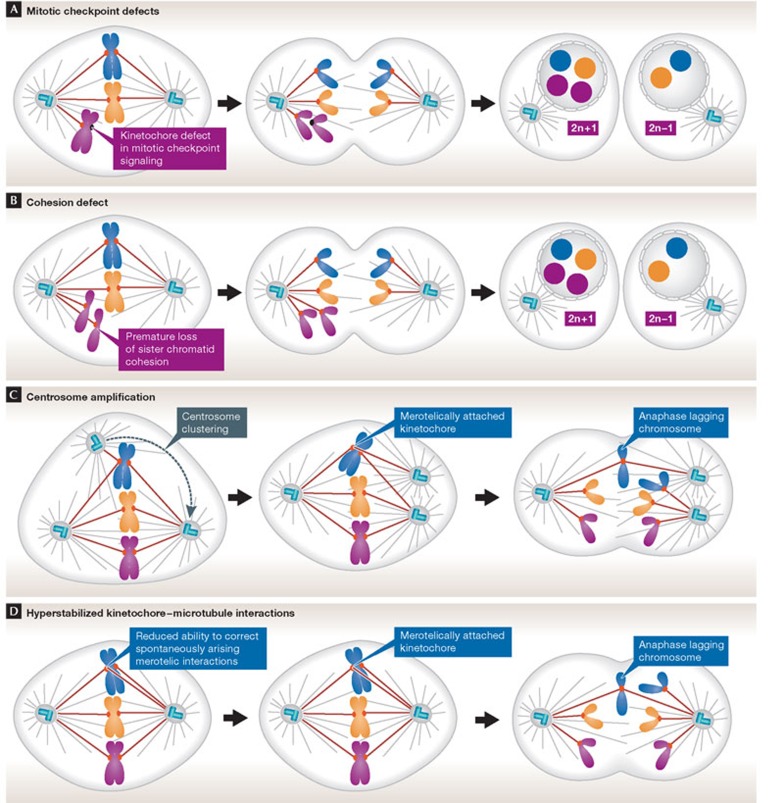
There are several pathways by which a cell might gain or lose chromosomes during mitosis. (A) Defects in mitotic checkpoint signalling. A compromised checkpoint allows onset of anaphase with unattached kinetochores, resulting in both copies of one chromosome being partitioned into the same daughter cell. (B) Chromosome cohesion defects. Chromosomes might be missegregated if sister chromatid cohesion is lost prematurely or persists during anaphase. (C) Multipolar mitotic spindle. Cells with extra centrosomes form multipolar mitotic spindles. In most instances, centrosomes cluster into two groups before anaphase. Centrosome clustering increases the frequency of merotelic attachments, in which a single kinetochore attaches to microtubules arising from both sides of the spindle. Merotelic attachments are sufficient to silence the mitotic checkpoint and, if not corrected before anaphase, merotelically attached chromosomes lag in the spindle midzone. Lagging chromosomes are either missegregated or excluded from both daughter nuclei forming a micronucleus (see Fig 3). (D) Hyperstable kinetochore–microtubule attachments. Correction of kinetochore–microtubule attachment errors requires the release of incorrectly attached microtubules. The slow turnover of kinetochore microtubules in CIN cells reduces their ability to correct spontaneous attachment errors, thereby increasing the frequency of merotelic attachments. CIN, chromosomal instability.
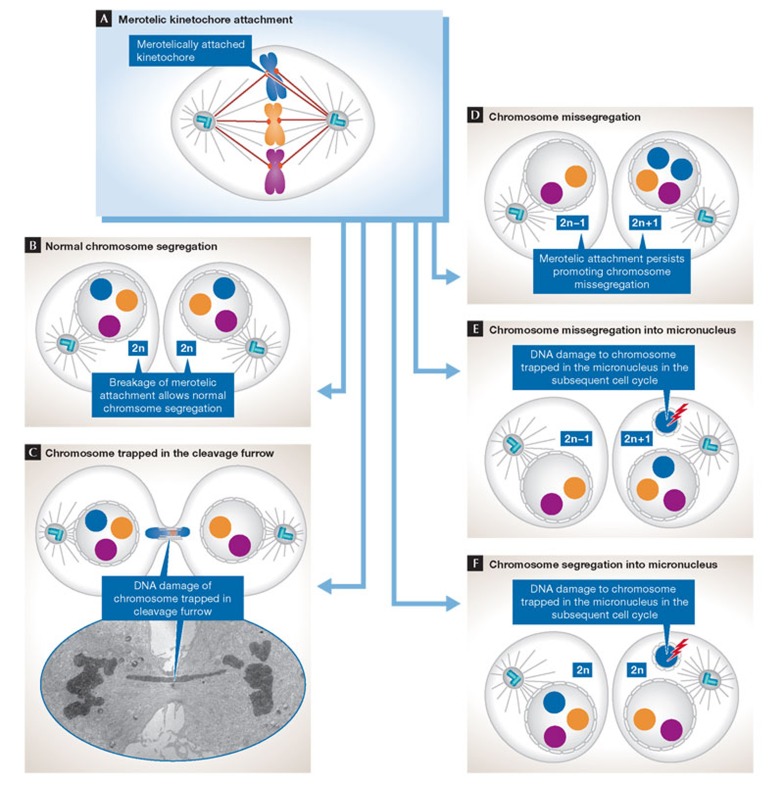
(A) Merotelic kinetochore–microtubule interactions arise when a single kinetochore is attached to microtubules emanating from both poles of the mitotic spindle. There are several possible fates for merotelically attached chromosomes. (B) Normal chromosome segregation. The merotelic kinetochore has only a few microtubules oriented to the wrong spindle pole and therefore the chromosome is segregated correctly. (C) The chromosome is trapped in the cleavage furrow. The merotelic kinetochore has approximately equal numbers of microtubules oriented towards the correct and incorrect pole, resulting in even pulling forces from both sides of the cell. The chromosome therefore remains in the centre of the cell and can be constricted by the cytokinetic furrow resulting in DNA damage that might cause subsequent chromosome rearrangements. The electron micrograph shows a chromosome lagging in the midzone of the spindle. (D) Chromosome missegregation. The merotelic kinetochore has many microtubules oriented to the wrong spindle pole and therefore the chromosome is segregated into the wrong daughter nucleus. (E) Chromosome missegregation into a micronucleus. The merotelic kinetochore lags in the middle of the spindle and is missegregated. In addition, the chromosome fails to incorporate into the daughter nucleus and forms a micronucleus. Chromosomes trapped in micronuclei accumulate high levels of DNA breaks in the subsequent cell cycle as a result of aberrant DNA replication or the cell entering mitosis while the micronuclei is still undergoing DNA replication. (F) Chromosome segregation into a micronucleus. The merotelic kinetochore lags in the middle of the spindle and is correctly segregated, but forms a micronucleus. As in (E), the micronucleus accumulates DNA damage in the next cell cycle that might lead to subsequent chromosome rearrangements.

Don W Cleveland & Andrew J Holland
References
-
- Hansemann D (1890) Ueber asymmetrische Zelltheilung in Epithelkrebsen und deren biologische Bedeutung. Arch Pathol Anat Physiol Klin Medicin 119: 299–326
-
- Boveri T (1914) Zur Frage der Entstenhung Maligner Tumoren. Jena, Germany: Gustav Fischer
-
- Mitelman F, Johansson B, Mertens FE (2012) Mitelman Database of Chromosome Aberrations and Gene Fusions in Cancer. http://cgap.nci.nih.gov/Chromosomes/Mitelman
-
- Paulsson K, Fioretos T, Strombeck B, Mauritzson N, Tanke HJ, Johansson B (2003) Trisomy 8 as the sole chromosomal aberration in myelocytic malignancies: a multicolor and locus-specific fluorescence in situ hybridization study. Cancer Genet Cytogenet 140: 66–69 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
