

Review
doi: 10.1038/embor.2012.54.
Age-related aneuploidy through cohesion exhaustion
Affiliations
- PMID: 22565322
- PMCID: PMC3367239
- DOI: 10.1038/embor.2012.54
Item in Clipboard
Review
Age-related aneuploidy through cohesion exhaustion
EMBO Rep.
.
Abstract
The trend of women to become pregnant when older than in previous generations poses a paramount medical problem, for oocytes are particularly prone to chromosome missegregation, and aneuploidy increases with age. Recent data strongly suggest that as oocyte age increases sister chromatid cohesion is weakened or lost. Cohesin deterioration seems to contribute significantly to age-dependent aneuploidy, as discussed in this review.
Conflict of interest statement
The author declares that he has no conflict of interest.
Figures
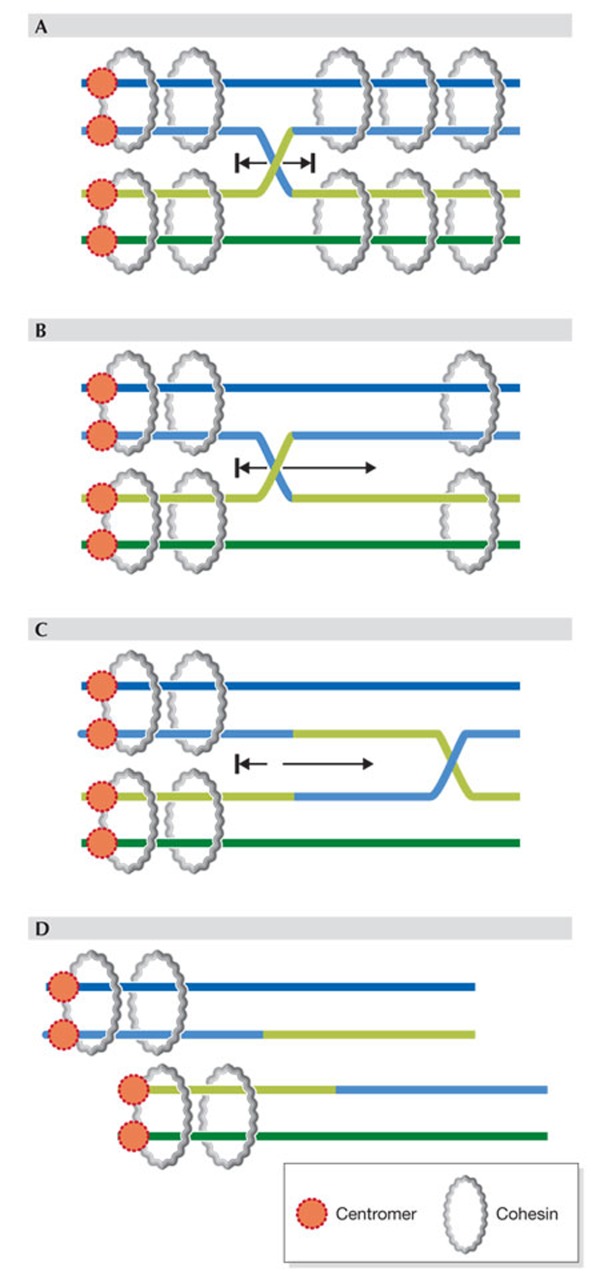
(A) Proper cohesion mediated by chromosome-wide distribution of cohesin rings prevents movement of cross-over or chiasma. (B–D) Loss of much arm cohesion allows movement of chiasma towards the centromere–distal end and eventually loss of chiasma (D), that is 'terminalization', whereby the two homologues (pairs of sister chromatids) separate prematurely.
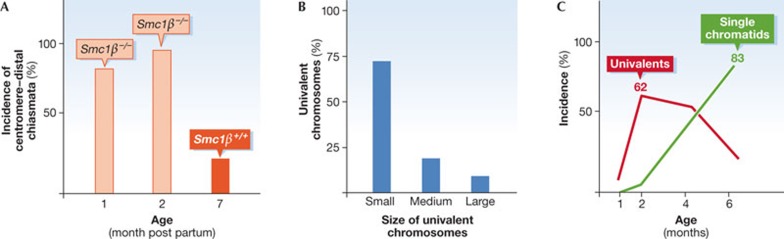
(A) Incidence of centromere–distal sites of chiasmata in oocytes from one-month-old and two-month-old female mice. Red columns: Smc1β−/− oocytes, yellow column: control value in Smc1β+/+ oocytes with low distal chiasmata even at seven months of age. (B) Distribution of metaphase I univalent chromosomes, which measures the loss of linkage of homologues, in Smc1β−/− oocytes depending on the length of the chromosomes, which are classified as short, medium or long. Short chromosomes suffer most loss of linkage, that is, loss of chiasmata. (C) Incidence of univalent chromosomes (red curve) and of single chromatids (green curve) with increasing age of the Smc1β−/− oocytes. Initially, homologues separate prematurely and univalents accumulate, which are later separated into single chromatids. In Smc1β+/+ oocytes there are no univalents at six months of age. For details see the work of Hodges and colleagues [44]. Smc1, structural maintenance of chromosomes.
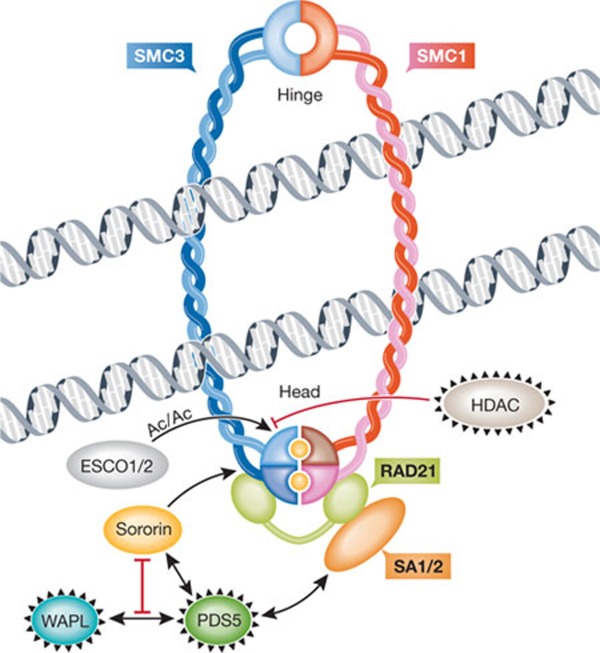
Anti-cohesion factors WAPL, HDAC and PDS5 are circled in black, and pro-cohesion factors are sororin and ESCO1/2. The hinge and head (amino/carboxy-terminal) domains of SMC proteins are indicated. Sister chromatids are shown being held together by a cohesin ring, that is, after cohesin has been loaded onto sister chromatids. The loading factors SCC2 and SCC4 are therefore not shown. Yellow circles within the cohesin head domains represent ATP, which needs to be bound and hydrolysed for stable cohesion. The relationship between ATP binding/hydrolysis and the action of pro- or anti-cohesion factors is unknown. The ring can open at the hinge domains, as postulated in the dual gate hypothesis [39], but is not depicted here as its regulation remains largely elusive. Other models of cohesin association with sister chromatids have been proposed, such as a 'handcuff' model comprising two cohesin rings that interact with each other [79]. AC, acetylation; ATP, adenosine triphosphate; ESCO, establishment of cohesion; HDAC, histone deacetylase; PDS5, precocious dissociation of sisters 5; SA, stromal antigen; SCC2/SCC4, sister chromatid cohesion 2/4; SMC1/3, structural maintenance of chromosomes; WAPL, wings apart-like.

Rolf Jessberger
References
-
- Hassold T, Hunt P (2001) To err (meiotically) is human: the genesis of human aneuploidy. Nat Rev Genet 2: 280–291 - PubMed
-
- Hassold T, Hall H, Hunt P (2007) The origin of human aneuploidy: where we have been, where we are going. Hum Mol Genet 16: R203–R208 - PubMed
-
- Homer H (2007) Ageing, aneuploidy and meiosis: eggs in a race against time. Yearbook of Obstetrics and Gynaecology 12: 139–158
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
