

Ranolazine decreases mechanosensitivity of the voltage-gated sodium ion channel Na(v)1.5: a novel mechanism of drug action
- PMID: 22565935
- PMCID: PMC3392778
- DOI: 10.1161/CIRCULATIONAHA.112.094714
Ranolazine decreases mechanosensitivity of the voltage-gated sodium ion channel Na(v)1.5: a novel mechanism of drug action
Abstract
Background: Na(V)1.5 is a mechanosensitive voltage-gated sodium-selective ion channel responsible for the depolarizing current and maintenance of the action potential plateau in the heart. Ranolazine is a Na(V)1.5 antagonist with antianginal and antiarrhythmic properties.
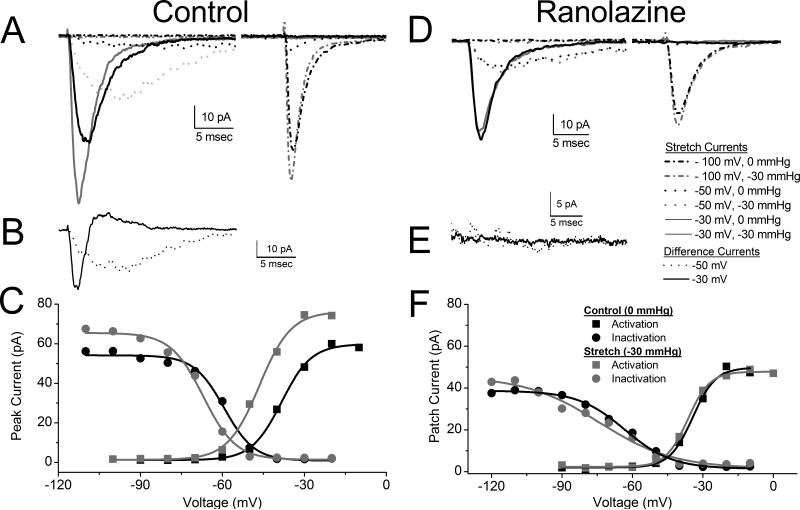
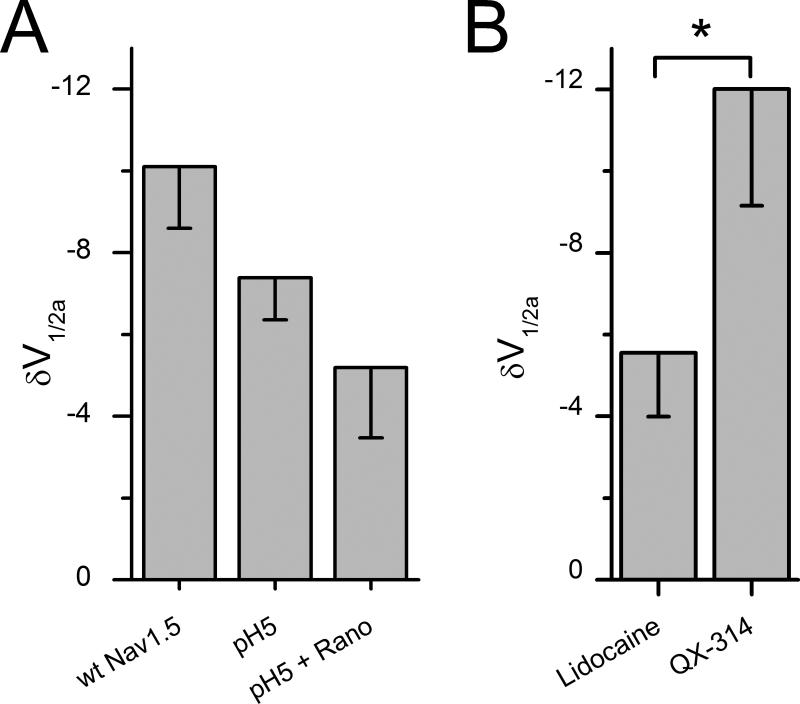
Methods and results: Mechanosensitivity of Na(V)1.5 was tested in voltage-clamped whole cells and cell-attached patches by bath flow and patch pressure, respectively. In whole cells, bath flow increased peak inward current in both murine ventricular cardiac myocytes (24±8%) and human embryonic kidney 293 cells heterologously expressing Na(V)1.5 (18±3%). The flow-induced increases in peak current were blocked by ranolazine. In cell-attached patches from cardiac myocytes and Na(V)1.5-expressing human embryonic kidney 293 cells, negative pressure increased Na(V) peak currents by 27±18% and 18±4% and hyperpolarized voltage dependence of activation by -11 mV and -10 mV, respectively. In human embryonic kidney 293 cells, negative pressure also increased the window current (250%) and increased late open channel events (250%). Ranolazine decreased pressure-induced shift in the voltage dependence (IC(50) 54 μmol/L) and eliminated the pressure-induced increases in window current and late current event numbers. Block of Na(V)1.5 mechanosensitivity by ranolazine was not due to the known binding site on DIVS6 (F1760). The effect of ranolazine on mechanosensitivity of Na(V)1.5 was approximated by lidocaine. However, ionized ranolazine and charged lidocaine analog (QX-314) failed to block mechanosensitivity.
Conclusions: Ranolazine effectively inhibits mechanosensitivity of Na(V)1.5. The block of Na(V)1.5 mechanosensitivity by ranolazine does not utilize the established binding site and may require bilayer partitioning. Ranolazine block of Na(V)1.5 mechanosensitivity may be relevant in disorders of mechanoelectric dysfunction.
Figures






Comment in
-
Cardiac sodium channel Na(v)1.5 mechanosensitivity is inhibited by ranolazine.Circulation. 2012 Jun 5;125(22):2681-3. doi: 10.1161/CIRCULATIONAHA.112.110908. Epub 2012 May 7. Circulation. 2012. PMID: 22565937 No abstract available.
References
-
- Cooper PJ, Lei M, Cheng LX, Kohl P. Selected contribution: Axial stretch increases spontaneous pacemaker activity in rabbit isolated sinoatrial node cells. J Appl Physiol. 2000;89:2099–2104. - PubMed
-
- Sung D, Mills RW, Schettler J, Narayan SM, Omens JH, McCulloch AD. Ventricular filling slows epicardial conduction and increases action potential duration in an optical mapping study of the isolated rabbit heart. J Cardiovasc Electrophysiol. 2003;14:739–749. - PubMed
-
- Maron BJ, Poliac LC, Kaplan JA, Mueller FO. Blunt impact to the chest leading to sudden death from cardiac arrest during sports activities. N Engl J Med. 1995;333:337–342. - PubMed
-
- Wang Z, Taylor LK, Denney WD, Hansen DE. Initiation of ventricular extrasystoles by myocardial stretch in chronically dilated and failing canine left ventricle. Circulation. 1994;90:2022–2031. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
