

In vivo T cell activation in lymphoid tissues is inhibited in the oxygen-poor microenvironment
- PMID: 22566817
- PMCID: PMC3342240
- DOI: 10.3389/fimmu.2011.00027
In vivo T cell activation in lymphoid tissues is inhibited in the oxygen-poor microenvironment
Abstract
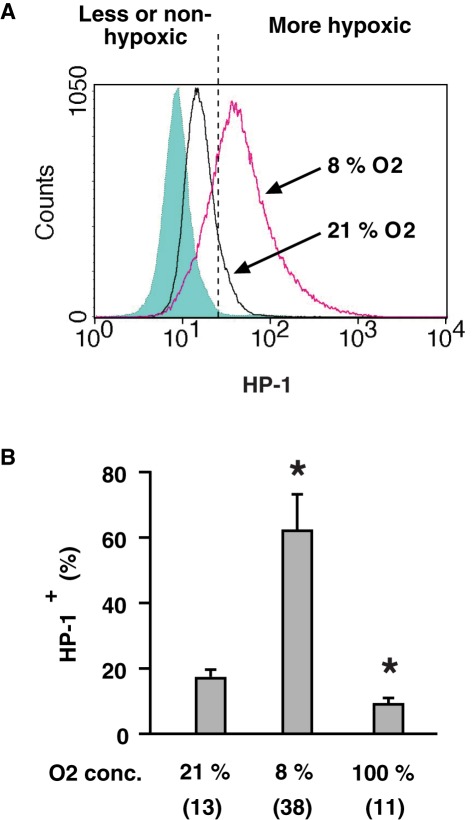
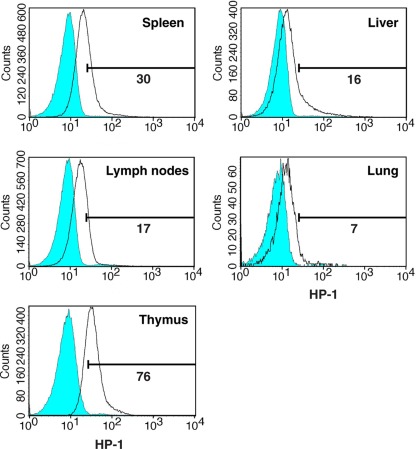
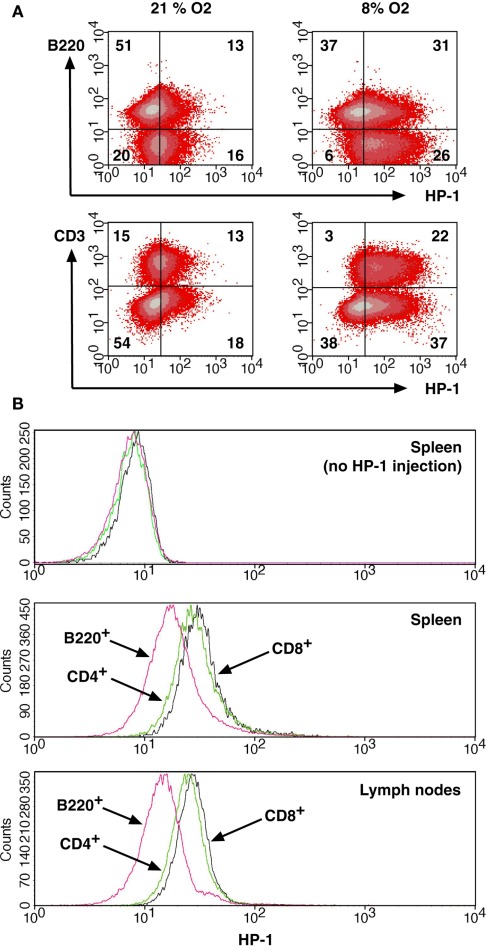
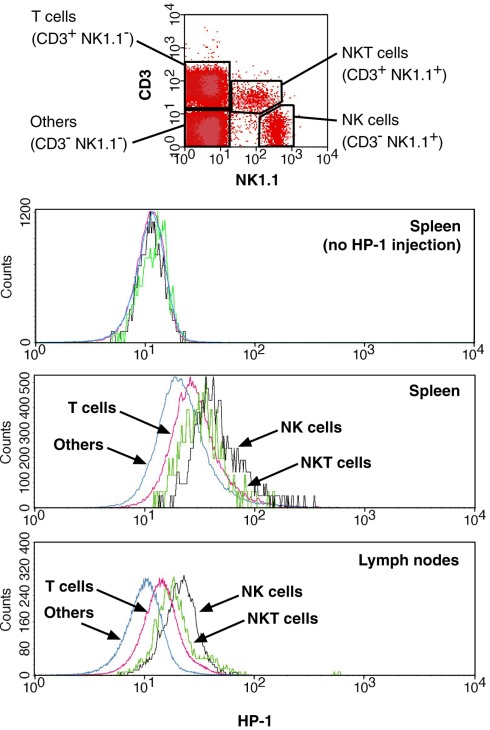
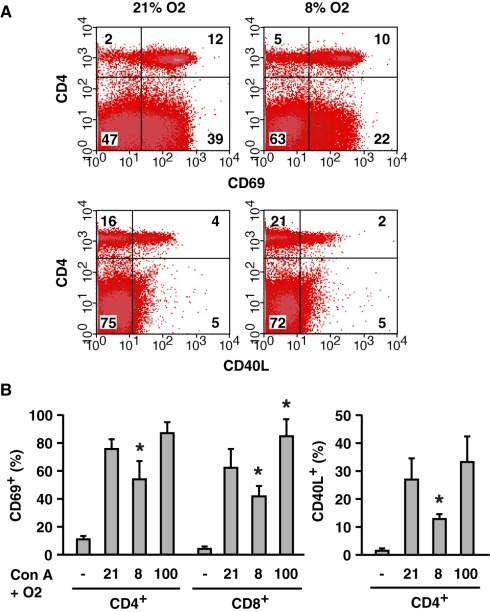
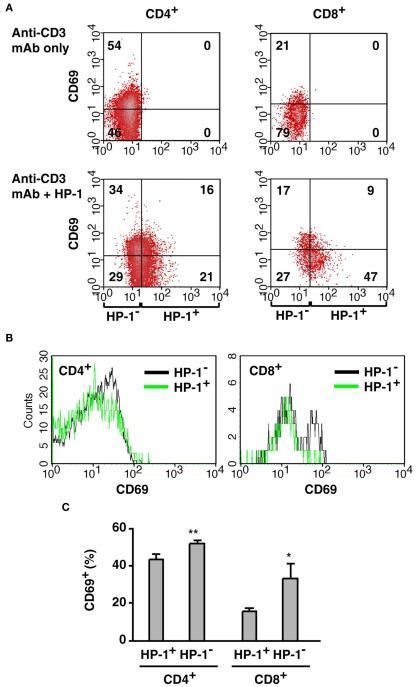
Activation of immune cells is under control of immunological and physiological regulatory mechanisms to ensure adequate destruction of pathogens with the minimum collateral damage to "innocent" bystander cells. The concept of physiological negative regulation of immune response has been advocated based on the finding of the critical immunoregulatory role of extracellular adenosine. Local tissue oxygen tension was proposed to function as one of such physiological regulatory mechanisms of immune responses. In the current study, we utilized in vivo marker of local tissue hypoxia pimonidazole hydrochloride (Hypoxyprobe-1) in the flowcytometric analysis of oxygen levels to which lymphocytes are exposed in vivo. The level of exposure to hypoxia in vivo was low in B cells and the levels increased in the following order: T cells < NKT cells < NK cells. The thymus was more hypoxic than the spleen and lymph nodes, suggesting the variation in the degree of oxygenation among lymphoid organs and cell types in normal mice. Based on in vitro studies, tissue hypoxia has been assumed to be suppressive to T cell activation in vivo, but there was no direct evidence demonstrating that T cells exposed to hypoxic environment in vivo are less activated. We tested whether the state of activation of T cells in vivo changes due to their exposure to hypoxic tissue microenvironments. The parallel analysis of more hypoxic and less hypoxic T cells in the same mouse revealed that the degree of T cell activation was significantly stronger in better-oxygenated T cells. These observations suggest that the extent of T cell activation in vivo is dependent on their localization and is decreased in environment with low oxygen tension.
Keywords: Hypoxyprobe-1; T cell; cytometry; hyperoxia; hypoxia; oxygen; tumor.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources