

The ambiguity in immunology
- PMID: 22566903
- PMCID: PMC3341998
- DOI: 10.3389/fimmu.2012.00018
The ambiguity in immunology
Abstract
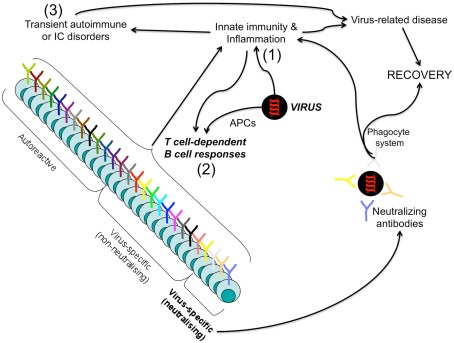
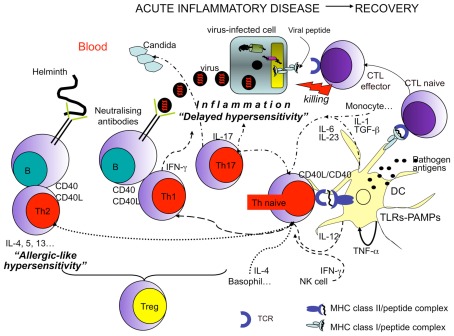
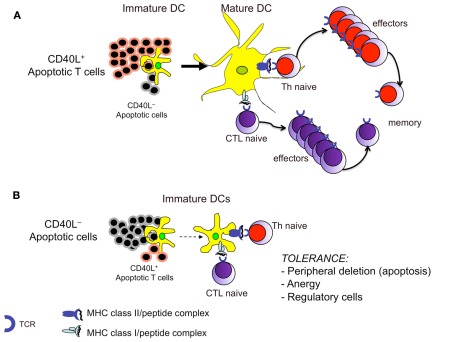
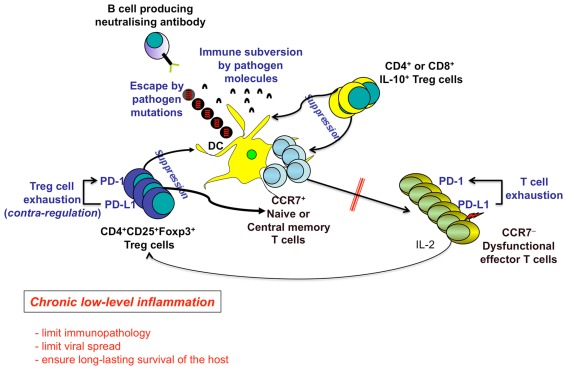
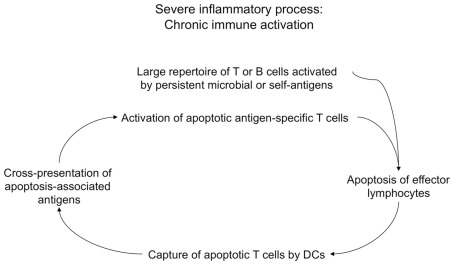
In the present article, we discuss the various ambiguous aspects of the immune system that render this complex biological network so highly flexible and able to defend the host from different external invaders. This ambiguity stems mainly from the property of the immune system to be both protective and harmful. Immunity cannot be fully protective without producing a certain degree of damage (immunopathology) to the host. The balance between protection and tissue damage is, therefore, critical for the establishment of immune homeostasis and protection. In this review, we will consider as ambiguous, various immunological tactics including: (a) the opposing functions driving immune responses, immune-regulation, and contra-regulation, as well as (b) the phenomenon of chronic immune activation as a result of a continuous cross-presentation of apoptotic T cells by dendritic cells. All these plans participate principally to maintain a state of chronic low-level inflammation during persisting infections, and ultimately to favor the species survival.
Keywords: ambiguity; autoimmunity; cancer; homeostasis; inflammation.
Figures

References
-
- Accapezzato D., Francavilla V., Paroli M., Casciaro M., Chircu L. V., Cividini A., Abrignani S., Mondelli M. U., Barnaba V. (2004). Hepatic expansion of a virus-specific regulatory CD8(+) T cell population in chronic hepatitis C virus infection. J. Clin. Invest. 113, 963–97210.1172/JCI200420515 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources