

Hip anatomy and ontogeny of lower limb musculature in three species of nonhuman primates
- PMID: 22567295
- PMCID: PMC3335645
- DOI: 10.1155/2011/580864
Hip anatomy and ontogeny of lower limb musculature in three species of nonhuman primates
Abstract
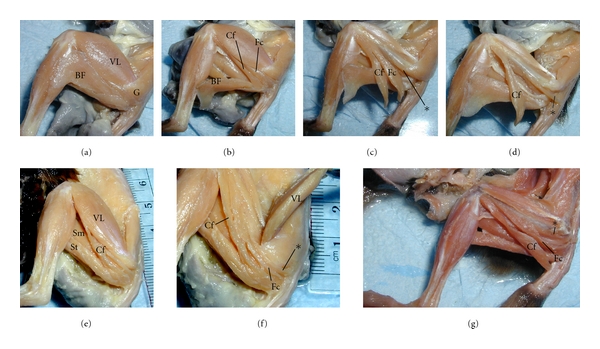
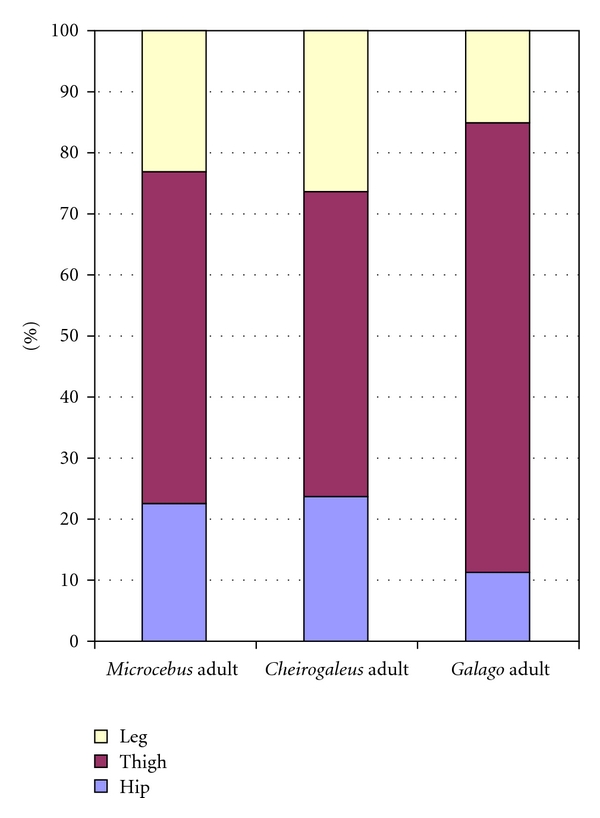
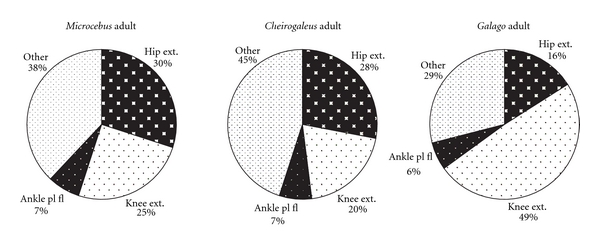
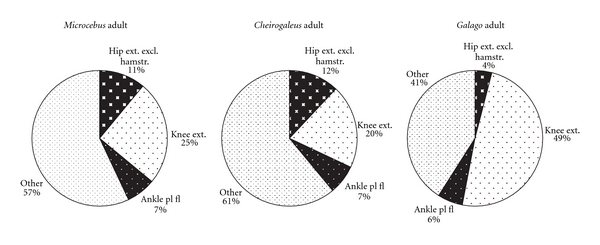
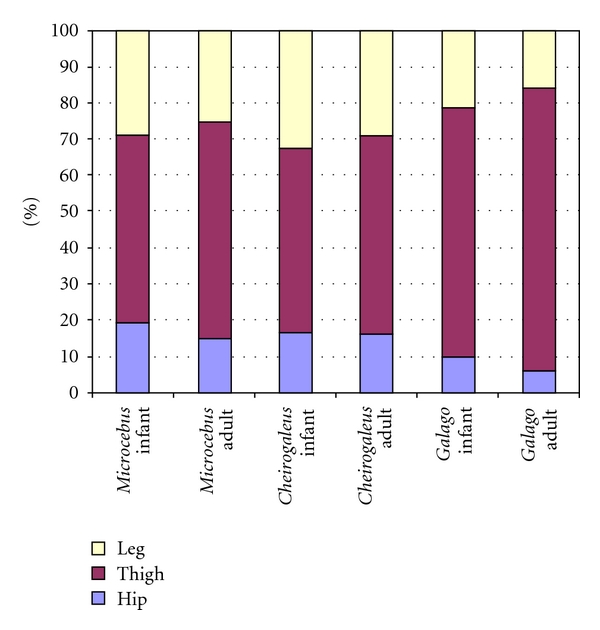
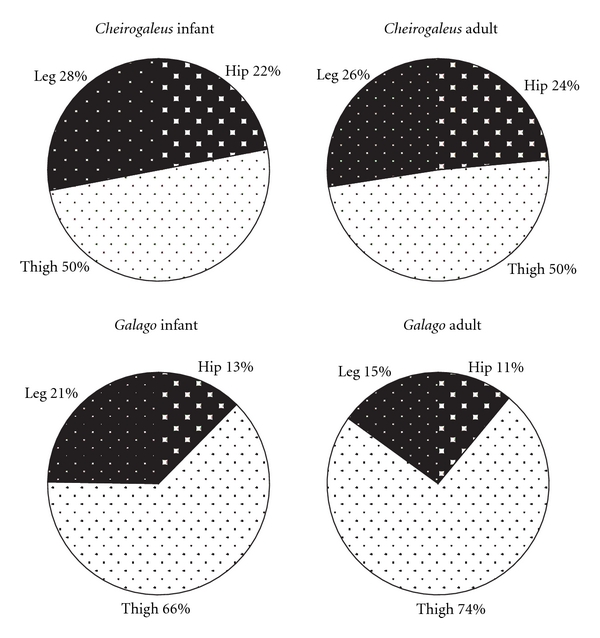
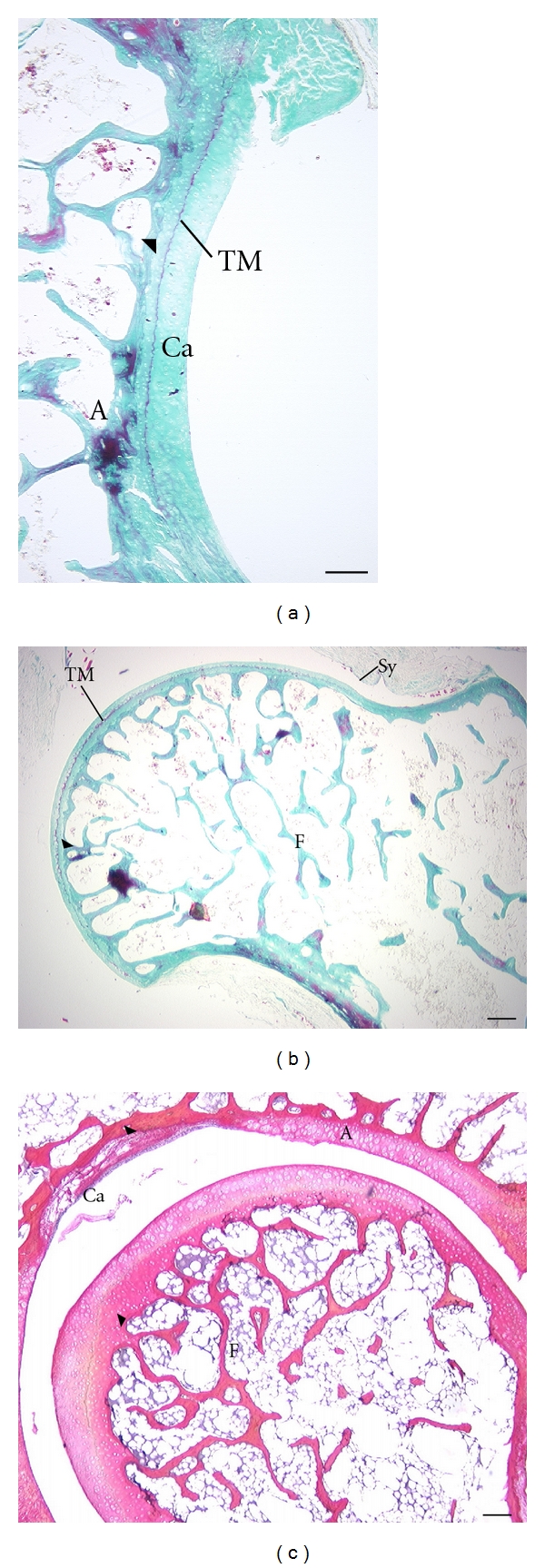
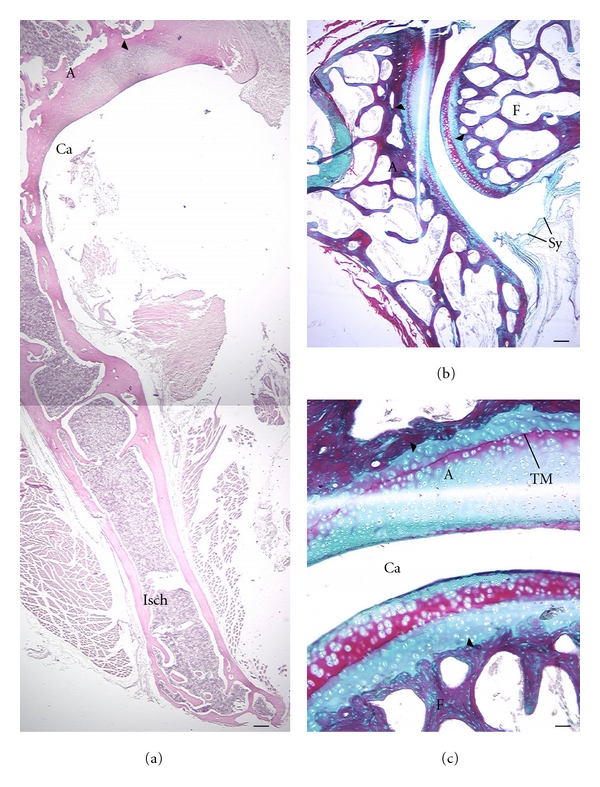
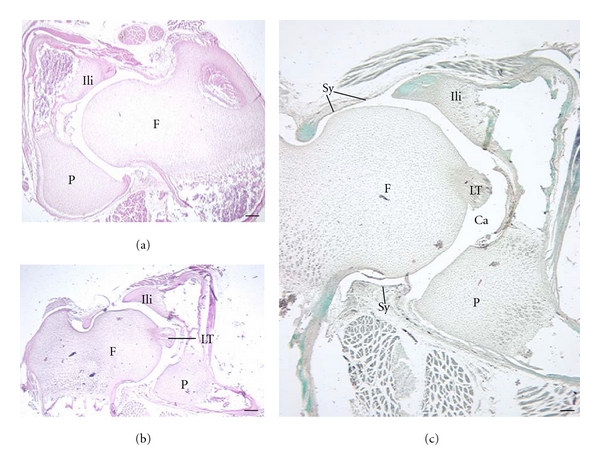
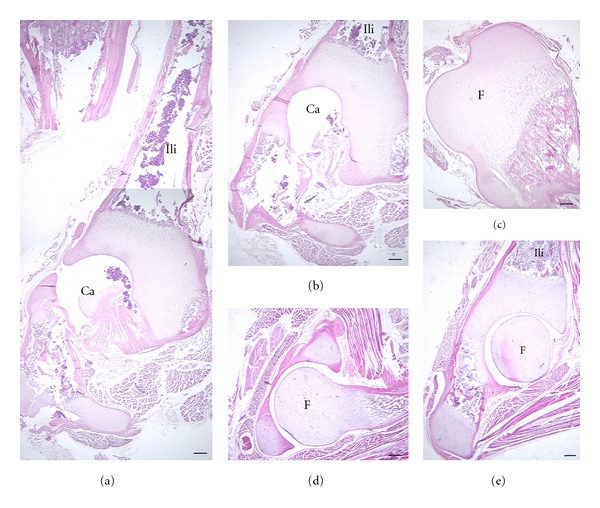
The hip region is examined to determine what aspects of musculoskeletal anatomy are precociously developed in primate species with highly specialized modes of locomotion. Muscles of the hind limb were removed and weighed in each specimen, and the hip joint of selected specimens was studied in stained serial sections. No perinatal differences among species are evident, but in adults, the hip joint of Galago moholi (a leaping specialist) appears to have proportionally thick articular cartilage (relative to the subchondral plate) compared to two species of cheirogaleids. Muscle mass distribution in the hind limbs confirms previous observations that the quadriceps femoris muscle is especially large in Galago (in percent mass of the entire hind limb), while the hip region is smaller compared to the more quadrupedal cheirogaleids. Across age groups, the species with the least specialized locomotion as adults, Cheirogaleus medius, shows little or no change in proximal to distal percentage distribution of muscle mass. Galago has a larger percentage mass gain in the thigh. We suggest that muscle mass gain to specific limb segments may be a critical milestone for primates with extremely specialized modes of locomotion.
Figures
References
-
- Atzeva M, Demes B, Kirkbride ML, Burrows AM, Smith TD. Comparison of hind limb muscle mass in neonate and adult prosimian primates. Journal of Human Evolution. 2007;52(3):231–242. - PubMed
-
- Anemone RL. The VCL hypothesis revisited: patterns of femoral morphology among quadrupedal and saltatorial prosimian primates. The American Journal of Physical Anthropology. 1990;83(3):373–393. - PubMed
-
- Anapol F, Barry K. Fiber architecture of the extensors of the hindlimb in semiterrestrial and arboreal guenons. The American Journal of Physical Anthropology. 1996;99(3):429–447. - PubMed
-
- Demes B, Fleagle JG, Lemelin P. Myological correlates of prosimian leaping. Journal of Human Evolution. 1998;34(4):385–399. - PubMed
-
- Anemone RL. The functional anatomy of the hip and thigh in primates. In: Gebo DL, editor. Postcranial Adaptation in Nonhuman Primates. DeKalb, Ill, USA: Northern Illinois University Press; 1993. pp. 151–174.
LinkOut - more resources
Full Text Sources
