

Principles of carbon catabolite repression in the rice blast fungus: Tps1, Nmr1-3, and a MATE-family pump regulate glucose metabolism during infection
- PMID: 22570632
- PMCID: PMC3342947
- DOI: 10.1371/journal.pgen.1002673
Principles of carbon catabolite repression in the rice blast fungus: Tps1, Nmr1-3, and a MATE-family pump regulate glucose metabolism during infection
Abstract
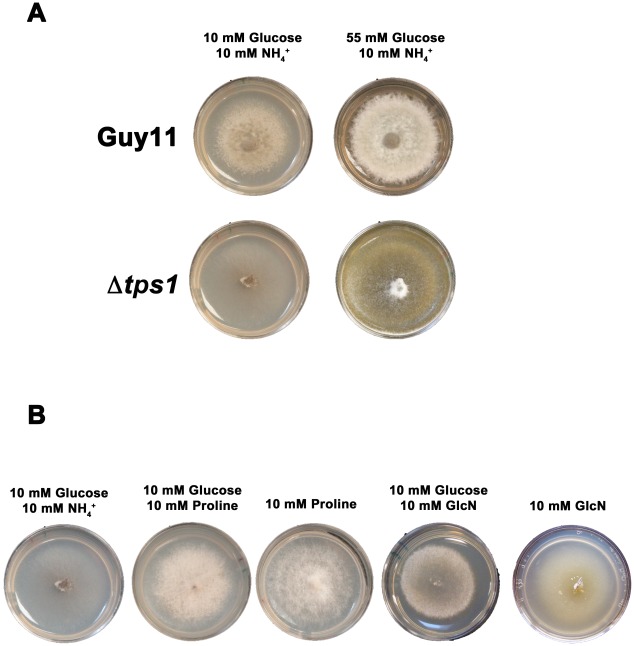
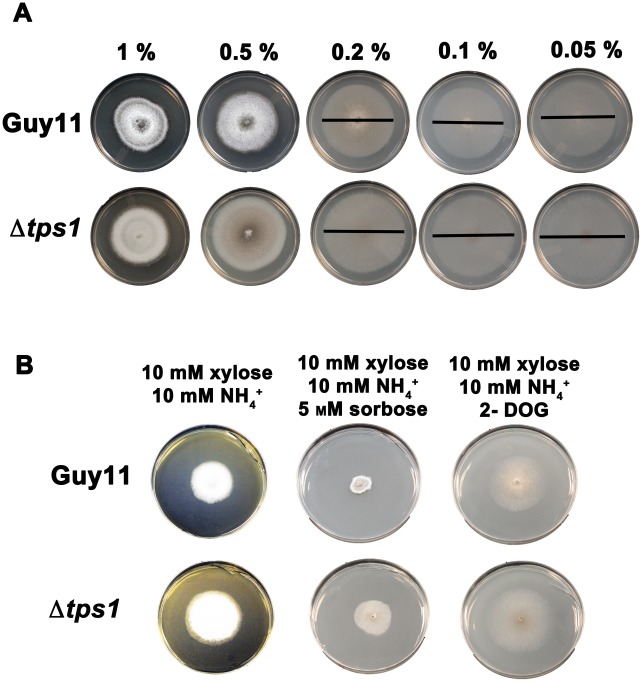
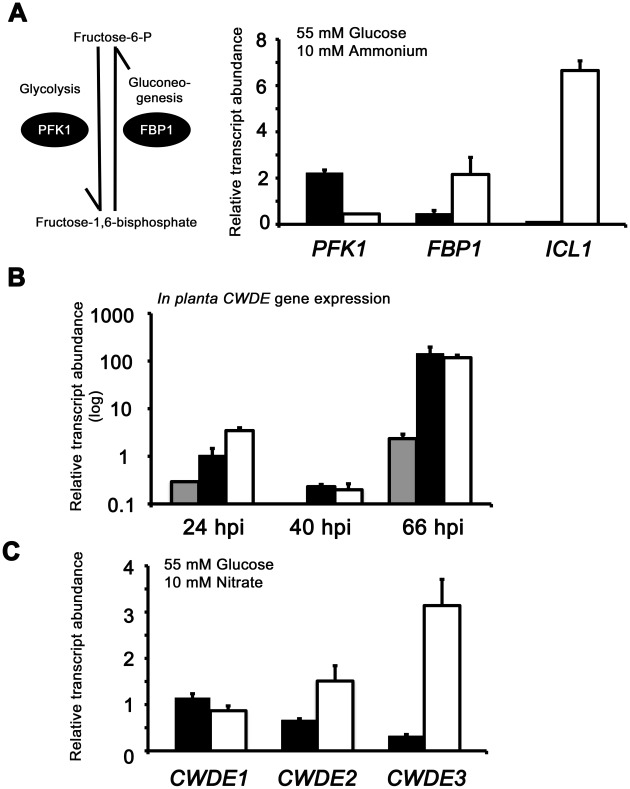
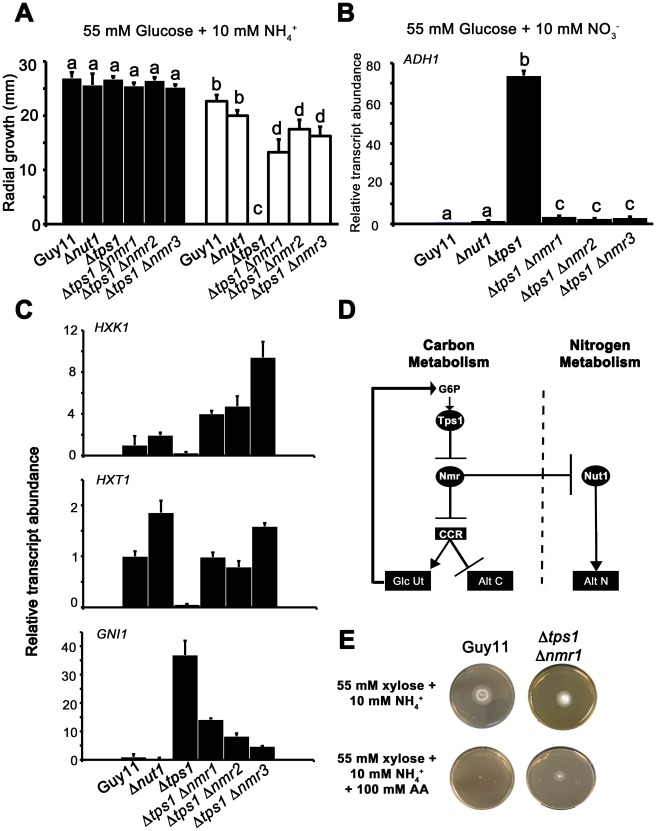
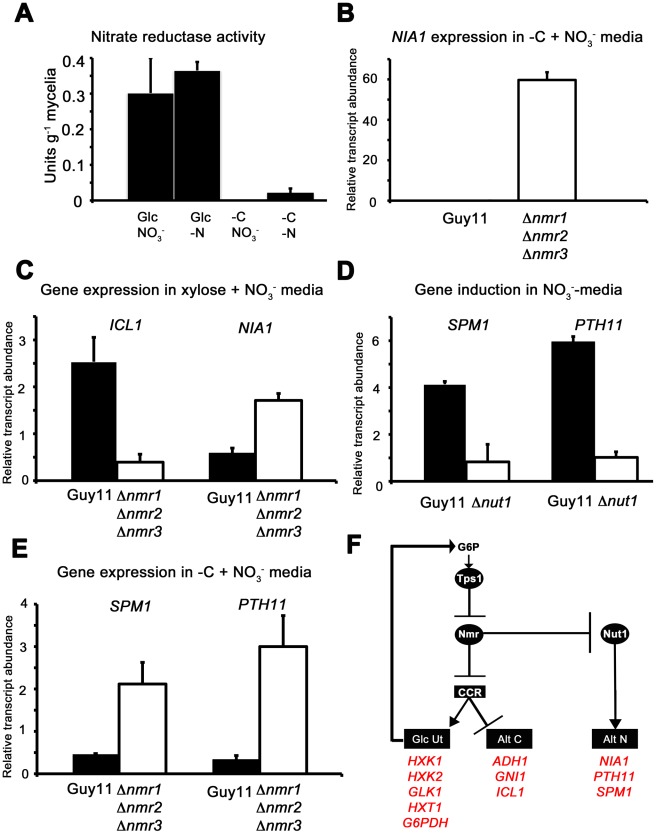
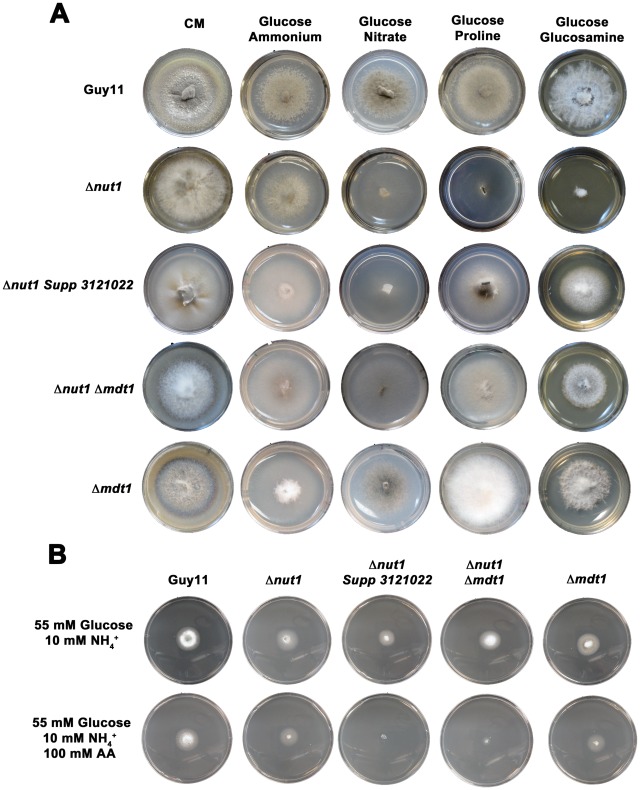
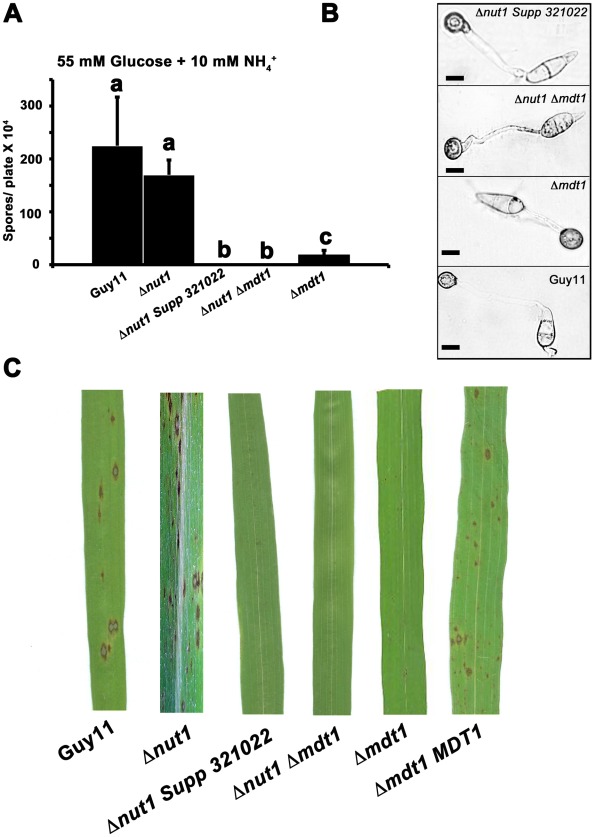
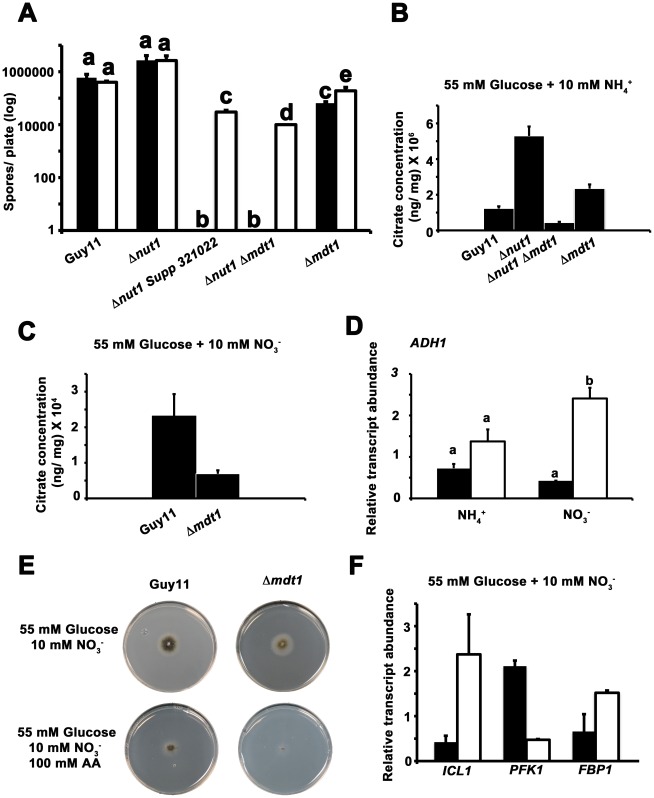
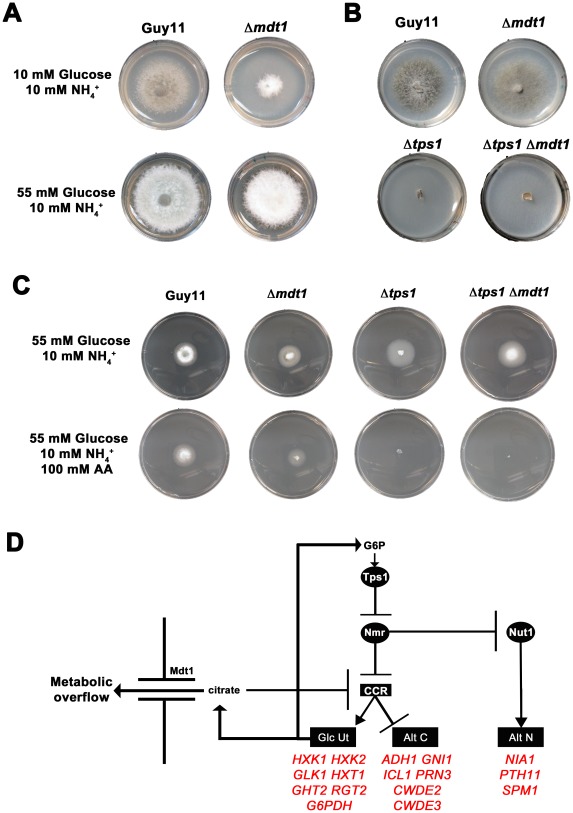
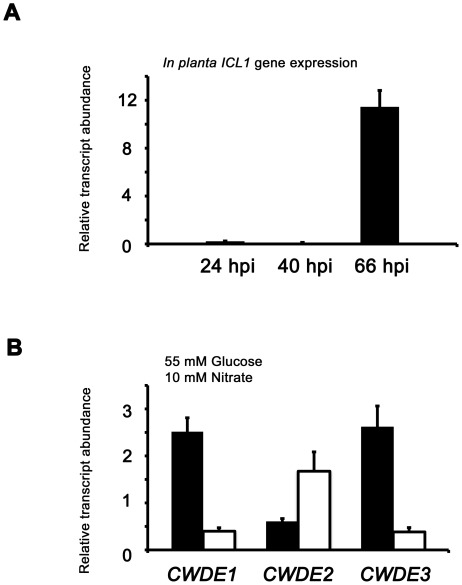
Understanding the genetic pathways that regulate how pathogenic fungi respond to their environment is paramount to developing effective mitigation strategies against disease. Carbon catabolite repression (CCR) is a global regulatory mechanism found in a wide range of microbial organisms that ensures the preferential utilization of glucose over less favourable carbon sources, but little is known about the components of CCR in filamentous fungi. Here we report three new mediators of CCR in the devastating rice blast fungus Magnaporthe oryzae: the sugar sensor Tps1, the Nmr1-3 inhibitor proteins, and the multidrug and toxin extrusion (MATE)-family pump, Mdt1. Using simple plate tests coupled with transcriptional analysis, we show that Tps1, in response to glucose-6-phosphate sensing, triggers CCR via the inactivation of Nmr1-3. In addition, by dissecting the CCR pathway using Agrobacterium tumefaciens-mediated mutagenesis, we also show that Mdt1 is an additional and previously unknown regulator of glucose metabolism. Mdt1 regulates glucose assimilation downstream of Tps1 and is necessary for nutrient utilization, sporulation, and pathogenicity. This is the first functional characterization of a MATE-family protein in filamentous fungi and the first description of a MATE protein in genetic regulation or plant pathogenicity. Perturbing CCR in Δtps1 and MDT1 disruption strains thus results in physiological defects that impact pathogenesis, possibly through the early expression of cell wall-degrading enzymes. Taken together, the importance of discovering three new regulators of carbon metabolism lies in understanding how M. oryzae and other pathogenic fungi respond to nutrient availability and control development during infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Arst HN, Cove DJ. Nitrogen metabolite repression in Aspergillus nidulans. Mol Gen Genet. 1973;126:111–141. - PubMed
-
- Bailey C, Arst HN. Carbon catabolite repression in Aspergillos nidulans. Eur J Biochem. 1975;51:573–577. - PubMed
-
- Caddick MX, Peters D, Platt A. Nitrogen regulation in fungi. Antonie Van Leeuwenhoek. 1994;65:169–177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
