

Transcriptional regulation by coactivators in embryonic stem cells
- PMID: 22572610
- PMCID: PMC3371141
- DOI: 10.1016/j.tcb.2012.04.002
Transcriptional regulation by coactivators in embryonic stem cells
Abstract
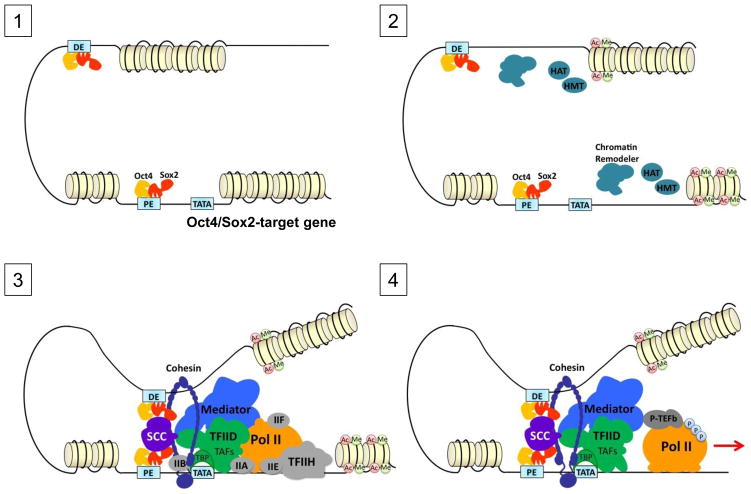
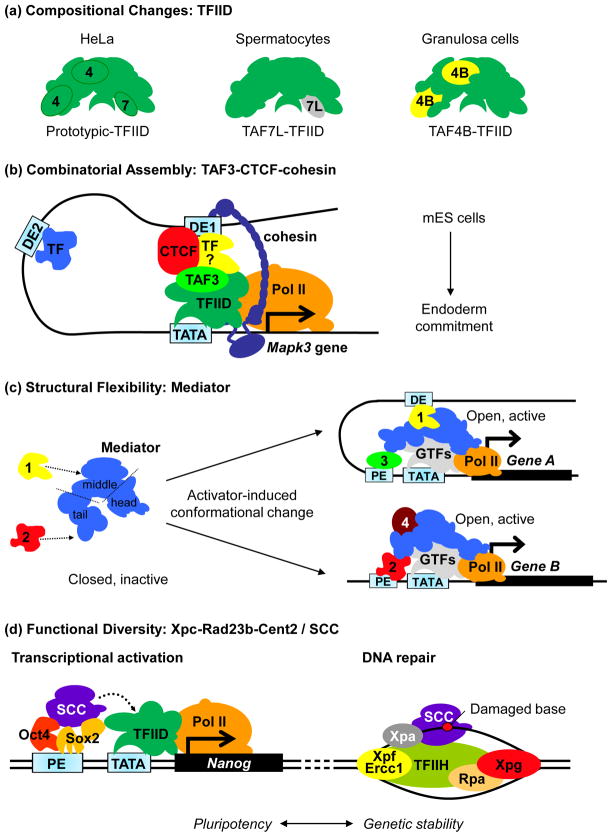
Embryonic stem (ES) cells, like all cell types, are defined by their unique transcriptional signatures. The ability of ES cells to self-renew or exit the pluripotent state and enter differentiation requires extensive changes in their transcriptome and epigenome. Remarkably, transcriptional programs governing each cell fate must remain sufficiently malleable so that expression of only a handful of transcriptional activators can override the pre-existing state by collaborating with an unexpectedly elaborate collection of coactivators to specify, restrict and stabilize the new state. Here, we discuss recent advances in our understanding of how the same coactivator can interpret multiple lines of information encoded by different activators and integrate signals from diverse regulators into stem cell-specific transcriptional outputs.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Transcriptional regulatory networks in embryonic stem cells.Cold Spring Harb Symp Quant Biol. 2008;73:203-9. doi: 10.1101/sqb.2008.73.026. Epub 2008 Nov 6. Cold Spring Harb Symp Quant Biol. 2008. PMID: 19022762 Review.
-
A core Klf circuitry regulates self-renewal of embryonic stem cells.Nat Cell Biol. 2008 Mar;10(3):353-60. doi: 10.1038/ncb1698. Epub 2008 Feb 10. Nat Cell Biol. 2008. PMID: 18264089
-
Signed weighted gene co-expression network analysis of transcriptional regulation in murine embryonic stem cells.BMC Genomics. 2009 Jul 20;10:327. doi: 10.1186/1471-2164-10-327. BMC Genomics. 2009. PMID: 19619308 Free PMC article.
-
Detailed characterization of the mouse embryonic stem cell transcriptome reveals novel genes and intergenic splicing associated with pluripotency.BMC Genomics. 2008 Apr 9;9:155. doi: 10.1186/1471-2164-9-155. BMC Genomics. 2008. PMID: 18400104 Free PMC article.
-
Regulation of embryonic stem cell self-renewal and differentiation by TGF-beta family signaling.Sci China Life Sci. 2010 Apr;53(4):497-503. doi: 10.1007/s11427-010-0096-2. Epub 2010 May 7. Sci China Life Sci. 2010. PMID: 20596917 Review.
Cited by
-
Low complexity domains, condensates, and stem cell pluripotency.World J Stem Cells. 2021 May 26;13(5):416-438. doi: 10.4252/wjsc.v13.i5.416. World J Stem Cells. 2021. PMID: 34136073 Free PMC article. Review.
-
Single-molecule tracking in live cells reveals distinct target-search strategies of transcription factors in the nucleus.Elife. 2014 Jun 12;3:e02230. doi: 10.7554/eLife.02230. Elife. 2014. PMID: 24925319 Free PMC article.
-
Autophagy and cell reprogramming.Cell Mol Life Sci. 2015 May;72(9):1699-713. doi: 10.1007/s00018-014-1829-3. Epub 2015 Jan 9. Cell Mol Life Sci. 2015. PMID: 25572296 Free PMC article. Review.
-
The extended pluripotency protein interactome and its links to reprogramming.Curr Opin Genet Dev. 2014 Oct;28:16-24. doi: 10.1016/j.gde.2014.08.003. Epub 2014 Aug 24. Curr Opin Genet Dev. 2014. PMID: 25173149 Free PMC article. Review.
-
The dyskerin ribonucleoprotein complex as an OCT4/SOX2 coactivator in embryonic stem cells.Elife. 2014 Nov 19;3:e03573. doi: 10.7554/eLife.03573. Elife. 2014. PMID: 25407680 Free PMC article.
References
-
- Chen X, et al. Integration of external signaling pathways with the core transcriptional network in embryonic stem cells. Cell. 2008;133:1106–1117. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
