

Structural tuning of the fluorescent protein iLOV for improved photostability
- PMID: 22573334
- PMCID: PMC3381190
- DOI: 10.1074/jbc.M111.318881
Structural tuning of the fluorescent protein iLOV for improved photostability
Abstract
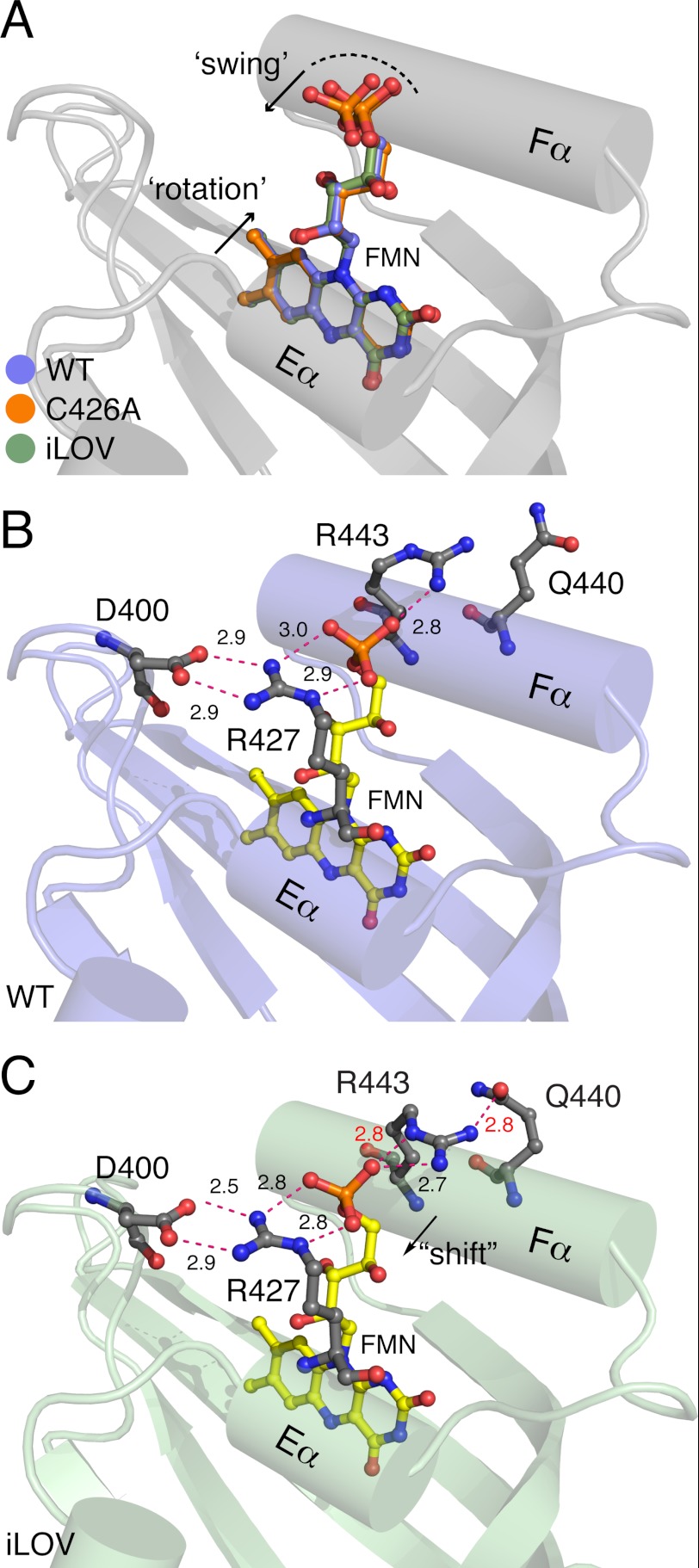
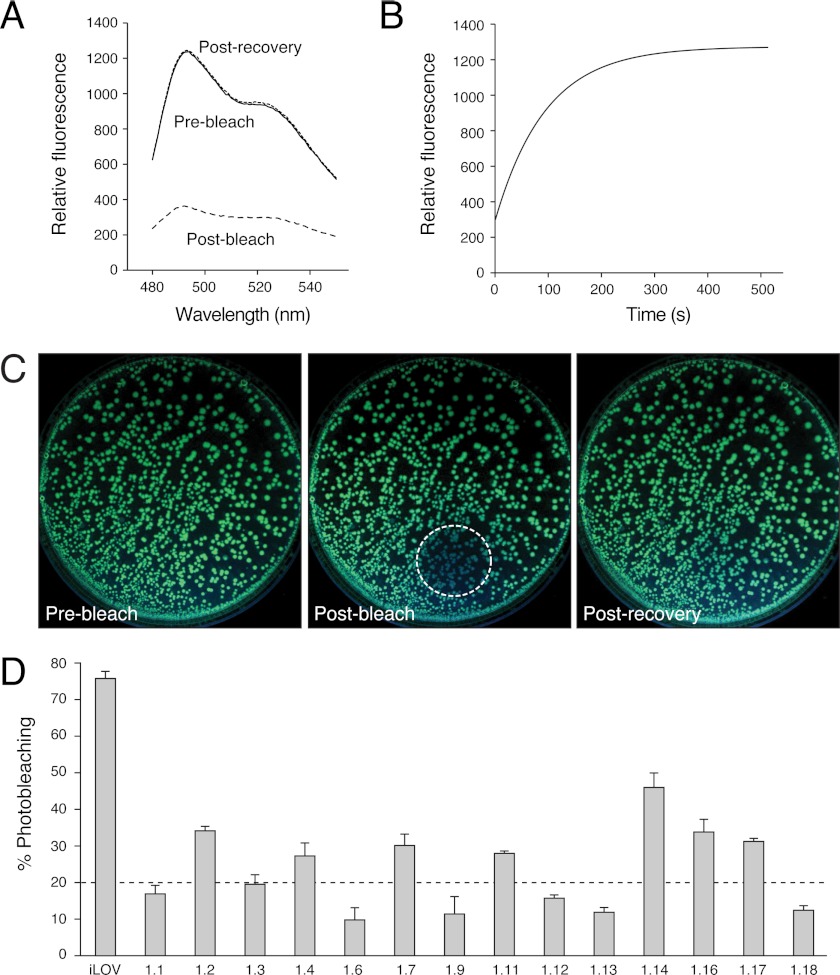
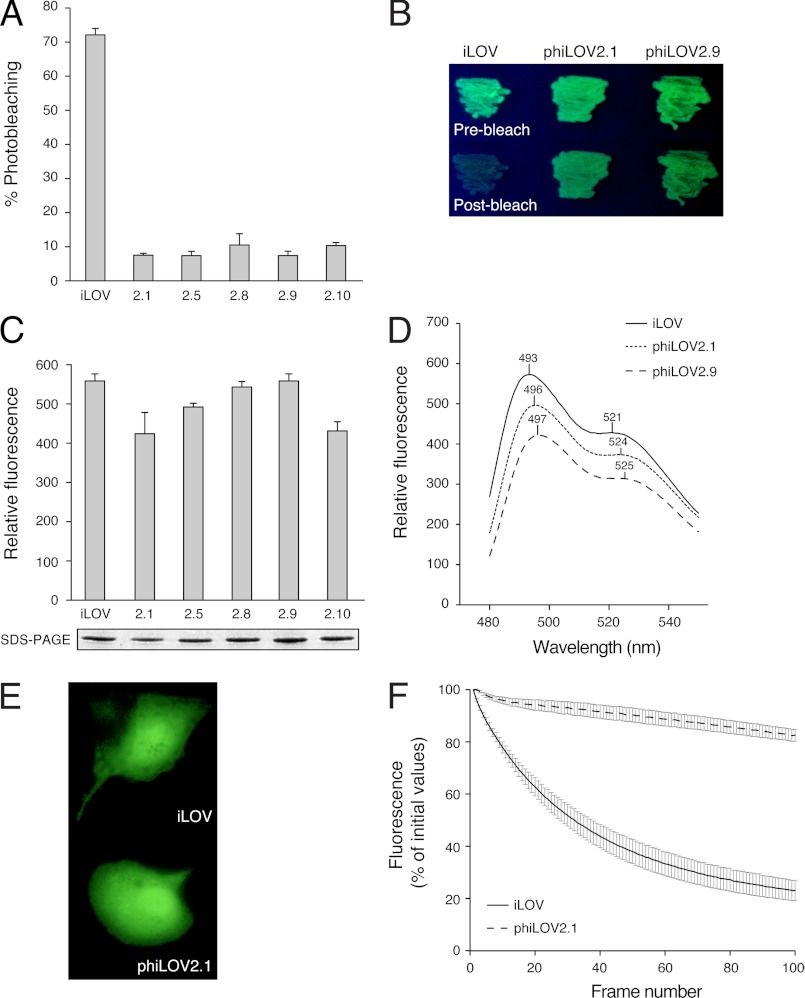
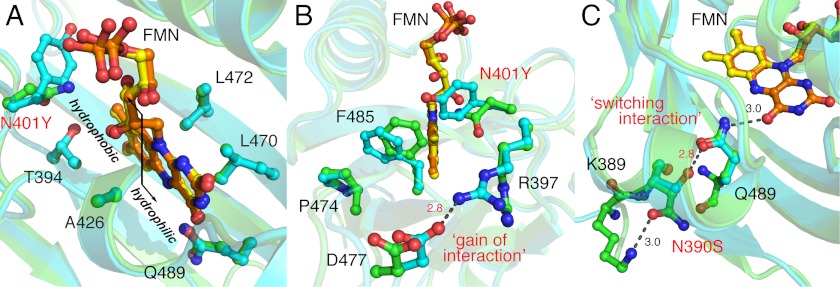
Fluorescent proteins derived from light, oxygen, or voltage (LOV) domains offer advantages over green fluorescent protein (GFP) from their small size and efficacy under anaerobic conditions. The flavoprotein improved LOV (iLOV) was engineered from the blue light receptor phototropin as a reporter of viral infection. To inform the molecular basis for the improved, photoreversible, fluorescent properties of iLOV, we employed directed evolution and determined five LOV crystallographic structures. Comparative structural analyses between iLOV and its progenitors reveal mutation-induced constraints in the environment of the flavin mononucleotide (FMN) chromophore; in iLOV, the methyl group of Thr-394 "crowds" the FMN isoalloxazine ring, Leu-470 triggers side chain "flipping" of Leu-472, and the terminal FMN phosphate shows increased anchoring. We further engineered iLOV variants that are readily detectable in bacterial and mammalian cells due to order-of-magnitude photostability increases. Structure determination of a resulting representative photostable iLOV (phiLOV) variant reveals additional constraints on the chromophore. Aromatic residues Tyr-401 and Phe-485 in phiLOV sandwich the FMN isoalloxazine ring from both sides, whereas Ser-390 anchors the side chain of FMN-interacting Gln-489 Our combined structural and mutational results reveal that constraining the FMN fluorophore yields improved photochemical properties for iLOV and its new photostable derivative. These findings provide a framework for structural fine-tuning of LOV scaffold proteins to maximize their potential as oxygen-independent fluorescent reporters.
Figures
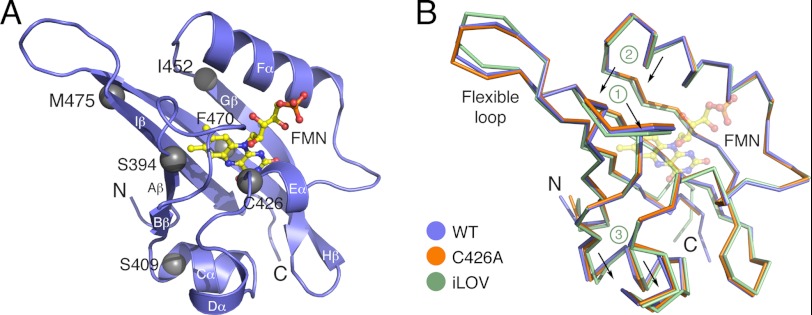
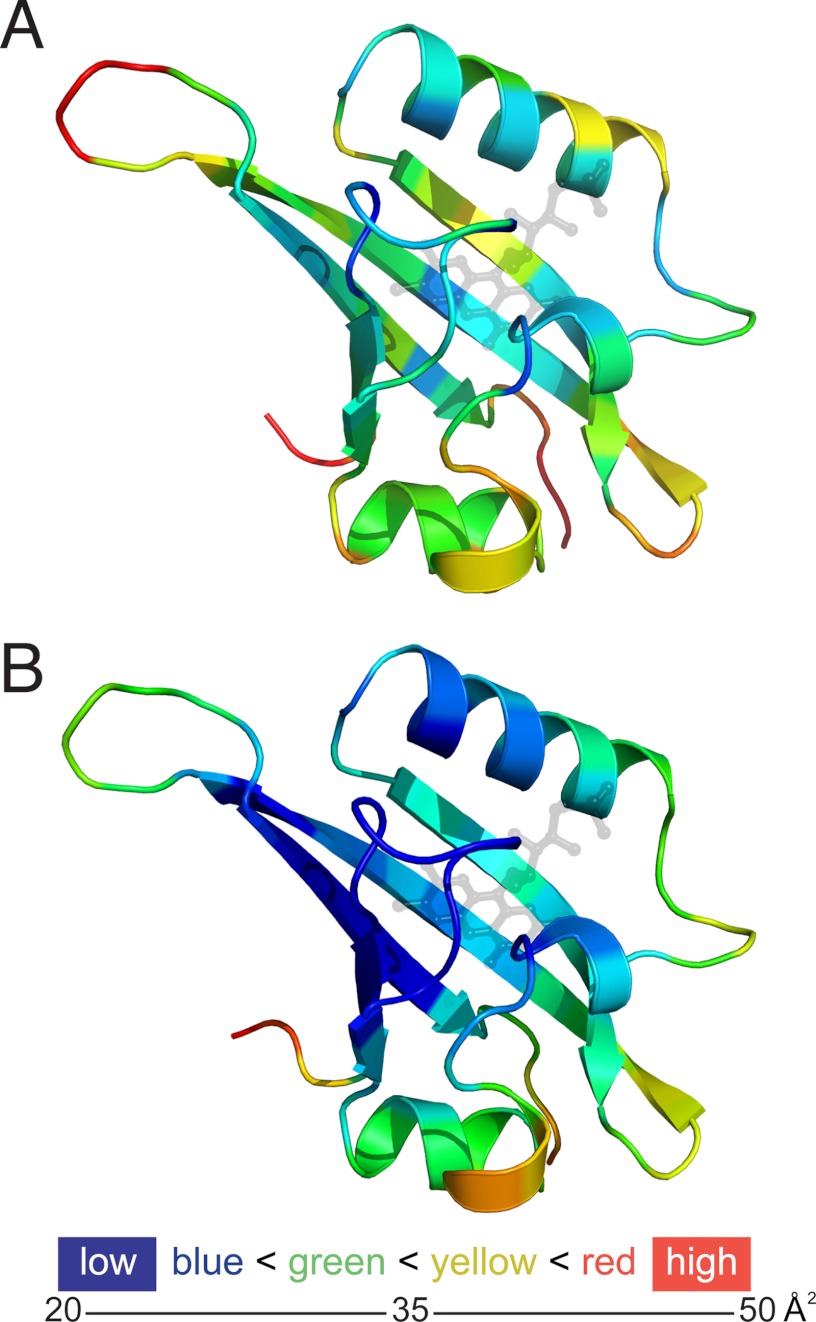
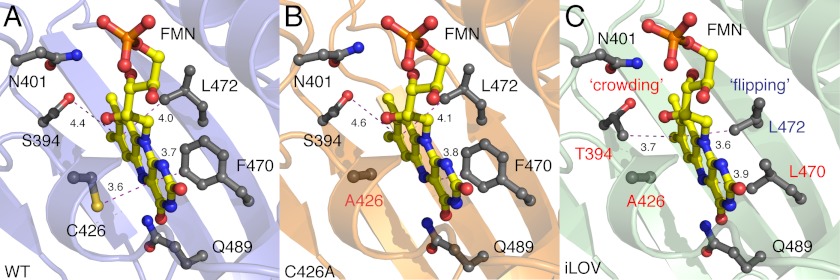
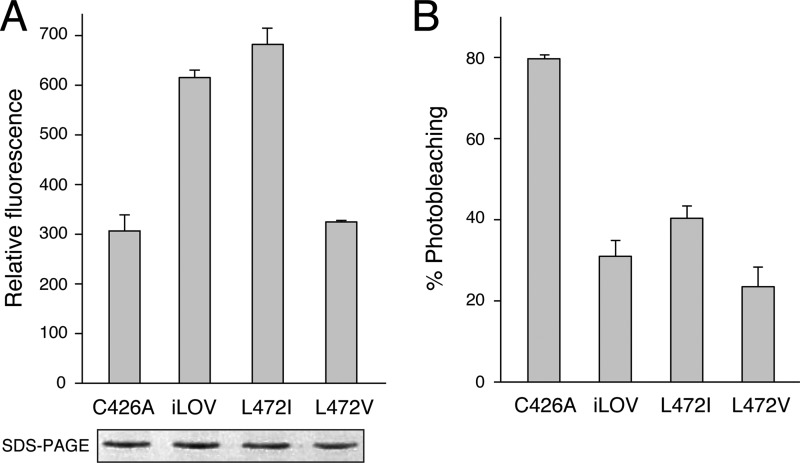
References
-
- Shaner N. C., Patterson G. H., Davidson M. W. (2007) Advances in fluorescent protein technology. J. Cell Sci. 120, 4247–4260 - PubMed
-
- Tsien R. Y. (1998) The green fluorescent protein. Annu. Rev. Biochem. 67, 509–544 - PubMed
-
- Drepper T., Eggert T., Circolone F., Heck A., Krauss U., Guterl J. K., Wendorff M., Losi A., Gärtner W., Jaeger K. E. (2007) Reporter proteins for in vivo fluorescence without oxygen. Nat. Biotechnol. 25, 443–445 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
