

Improving the performance of true single molecule sequencing for ancient DNA
- PMID: 22574620
- PMCID: PMC3430569
- DOI: 10.1186/1471-2164-13-177
Improving the performance of true single molecule sequencing for ancient DNA
Abstract
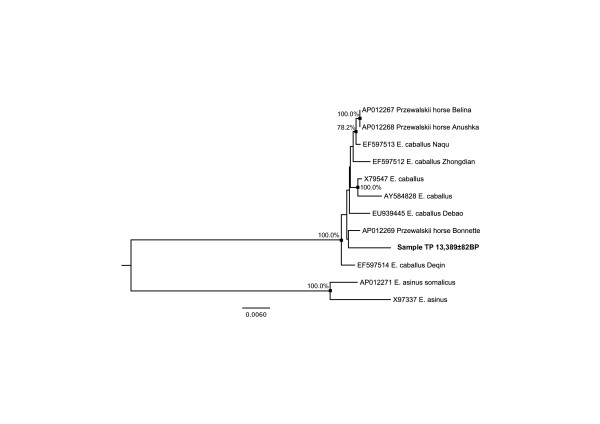
Background: Second-generation sequencing technologies have revolutionized our ability to recover genetic information from the past, allowing the characterization of the first complete genomes from past individuals and extinct species. Recently, third generation Helicos sequencing platforms, which perform true Single-Molecule DNA Sequencing (tSMS), have shown great potential for sequencing DNA molecules from Pleistocene fossils. Here, we aim at improving even further the performance of tSMS for ancient DNA by testing two novel tSMS template preparation methods for Pleistocene bone fossils, namely oligonucleotide spiking and treatment with DNA phosphatase.
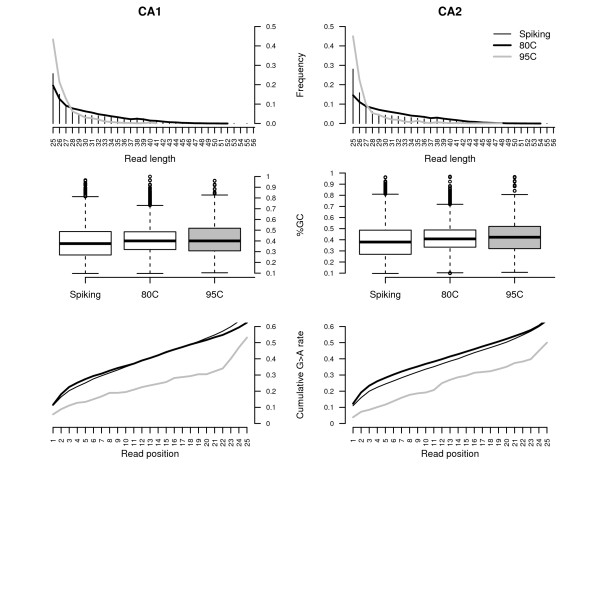
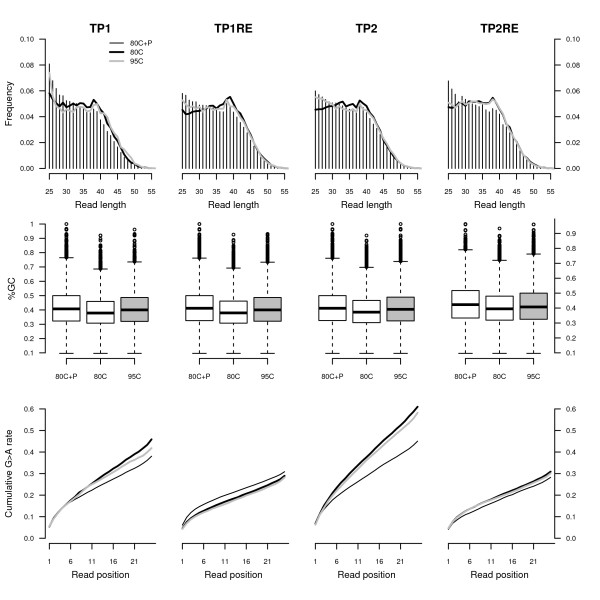
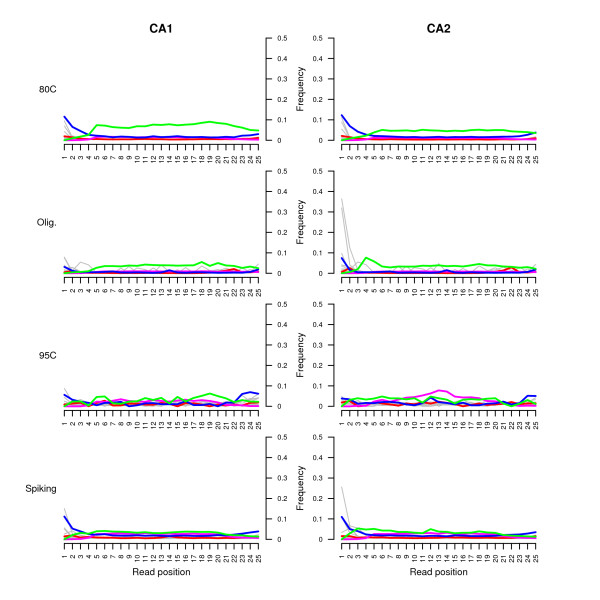
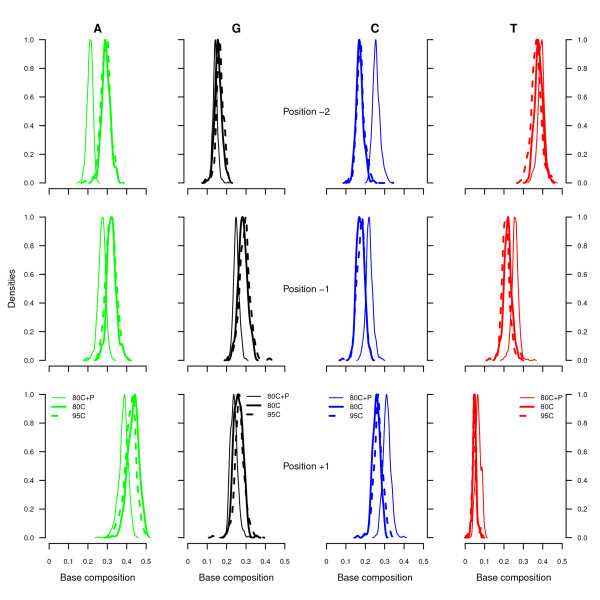
Results: We found that a significantly larger fraction of the horse genome could be covered following oligonucleotide spiking however not reproducibly and at the cost of extra post-sequencing filtering procedures and skewed %GC content. In contrast, we showed that treating ancient DNA extracts with DNA phosphatase improved the amount of endogenous sequence information recovered per sequencing channel by up to 3.3-fold, while still providing molecular signatures of endogenous ancient DNA damage, including cytosine deamination and fragmentation by depurination. Additionally, we confirmed the existence of molecular preservation niches in large bone crystals from which DNA could be preferentially extracted.
Conclusions: We propose DNA phosphatase treatment as a mechanism to increase sequence coverage of ancient genomes when using Helicos tSMS as a sequencing platform. Together with mild denaturation temperatures that favor access to endogenous ancient templates over modern DNA contaminants, this simple preparation procedure can improve overall Helicos tSMS performance when damaged DNA templates are targeted.
Figures
References
-
- Willerslev E, Cappellini E, Boosma W, Nielsen R, Hebsgaard MB, Brand TB, Hofreiter M, Bunce M, Poinar HN, Dahl-Jensen D, Johnsen S, Steffensen JP, Bennike O, Schwenninger JL, Nathan R, Armitage S, de Hoog CJ, Alfimov V, Christl M, Beer J, Muscheler R, Barker J, Sharp M, Penkman KE, Haile J, Taberlet P, Gilbert MT, Casoli A, Campani E, Collins MJ. Ancient biomolecules from deep ice cores reveal a forested southern Greenland. Science. 2007;317:111–114. doi: 10.1126/science.1141758. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
