

Safety study of adeno-associated virus serotype 2-mediated human acid sphingomyelinase expression in the nonhuman primate brain
- PMID: 22574943
- PMCID: PMC3413900
- DOI: 10.1089/hum.2012.052
Safety study of adeno-associated virus serotype 2-mediated human acid sphingomyelinase expression in the nonhuman primate brain
Abstract
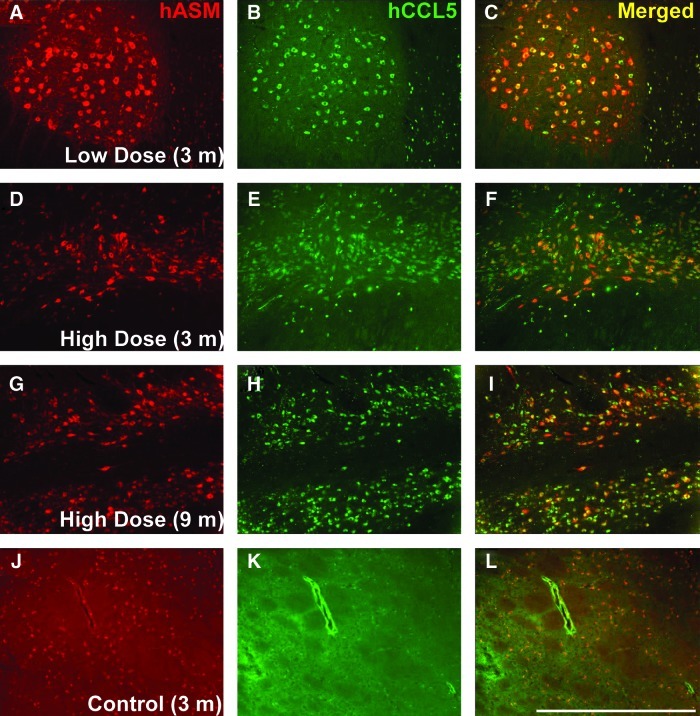
Niemann-Pick disease is a lysosomal storage disorder resulting from inherited deficiency in acid sphingomyelinase (ASM). Use of adeno-associated virus serotype 2 (AAV2) to deliver human acid sphingomyelinase (hASM) is currently being explored as a means to treat the devastating neurological features of NPD, which are refractory to traditional enzyme replacement therapy. In this study, we evaluated the long-term efficacy and safety of AAV2-hASM after direct infusion into the CNS of nonhuman primates. First, we confirmed the efficacy of AAV2-hASM in naive rats, which exhibited increased ASM expression and enzyme activity after infusion, without evidence of local or systemic toxicity. Next, the model was adapted to naive nonhuman primates (NHPs) with various doses of AAV2-hASM or saline delivered into the brainstem and both thalami. Strikingly, NHPs that received a high dose of AAV2-hASM displayed significant motor deficits that were not seen in low-dose animals in both the short-term (3-month) and long-term (9-month) treatment groups. In treated NHPs, ASM expression and activity were elevated with associated alterations in the sphingolipidomic profile in brain regions transduced with AAV2-hASM. Initial histological analysis indicated marked inflammatory reactions, and immunohistochemical analysis confirmed a robust inflammatory response. Importantly, pronounced upregulation of the chemokine CCL5, a target of ASM-mediated inflammatory signaling, was detected that correlated with the inflammatory response, providing a possible mechanism for hASM-associated toxicity. This study defines dose-dependent and dose-independent toxicities of AAV2-hASM in the naive primate brain, and reveals potential challenges in the design of a clinical trial.
Figures
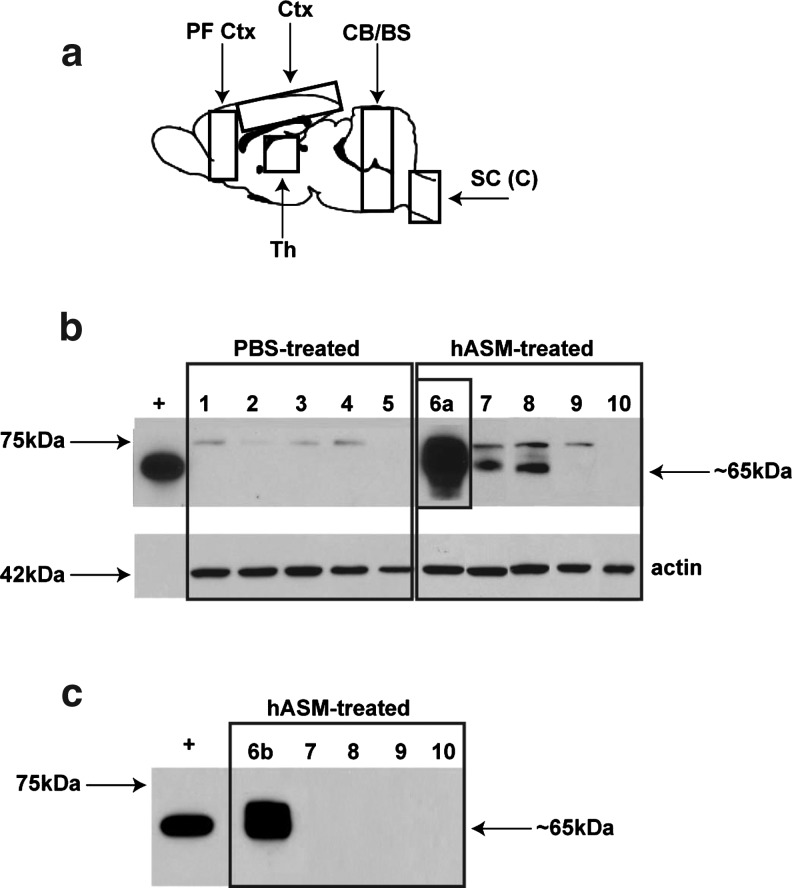

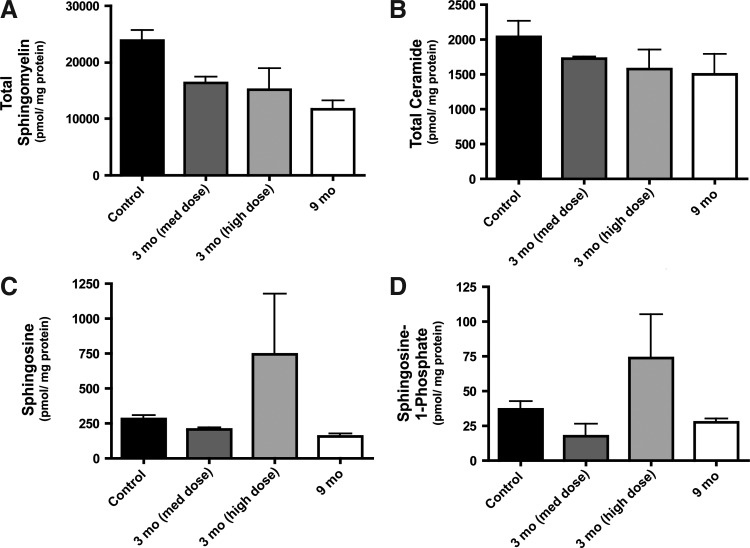
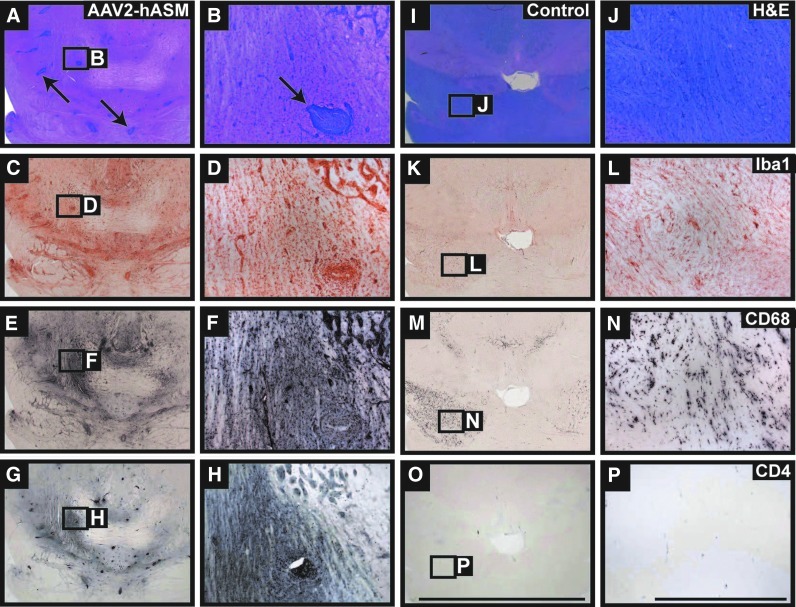
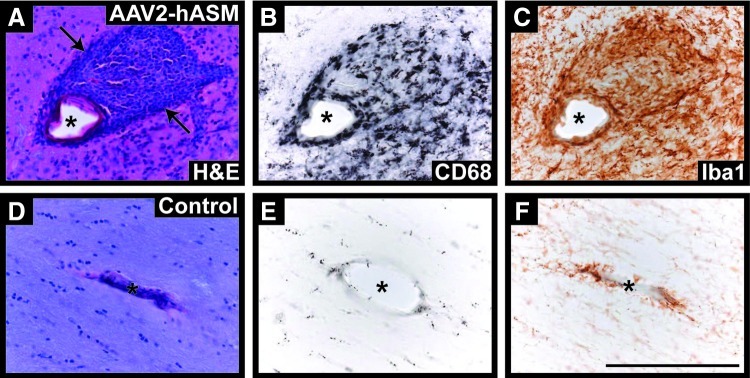
References
-
- Ayuso E. Mingozzi F. Montane J., et al. High AAV vector purity results in serotype- and tissue-independent enhancement of transduction efficiency. Gene Ther. 2010;17:503–510. - PubMed
-
- Barranger J.M. Novelli E.A. Gene therapy for lysosomal storage disorders. Expert Opin. Biol. Ther. 2001;1:857–867. - PubMed
-
- Bielawski J. Szulc Z.M. Hannun Y.A., et al. Simultaneous quantitative analysis of bioactive sphingolipids by high-performance liquid chromatography-tandem mass spectrometry. Methods. 2006;39:82–91. - PubMed
-
- Bosch A. Perret E. Desmaris N., et al. Reversal of pathology in the entire brain of mucopolysaccharidosis type VII mice after lentivirus-mediated gene transfer. Hum. Gene Ther. 2000;11:1139–1150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
