

Cell size control in bacteria
- PMID: 22575476
- PMCID: PMC3350639
- DOI: 10.1016/j.cub.2012.02.032
Cell size control in bacteria
Abstract
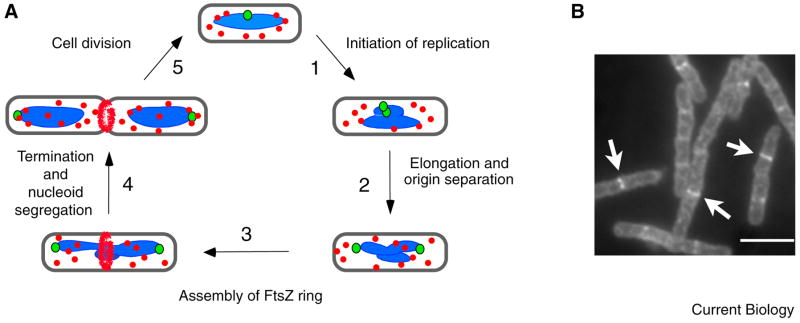
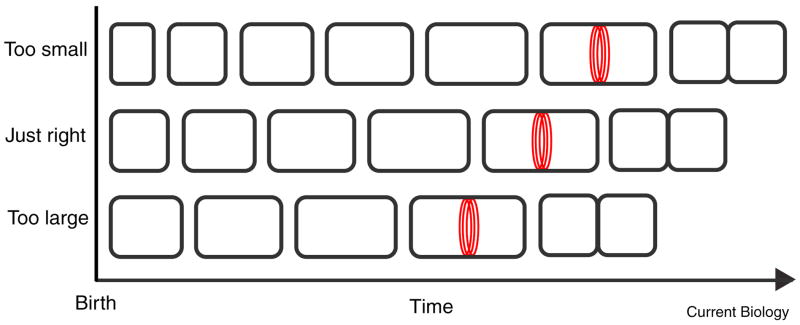
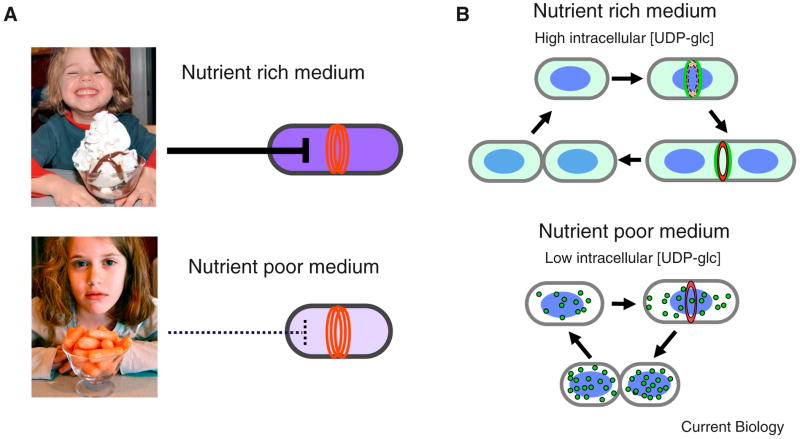
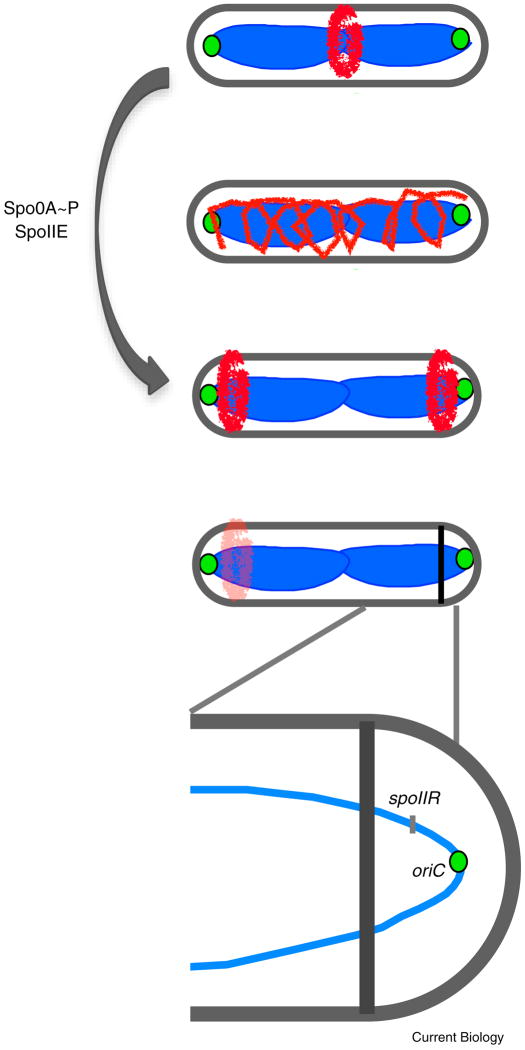
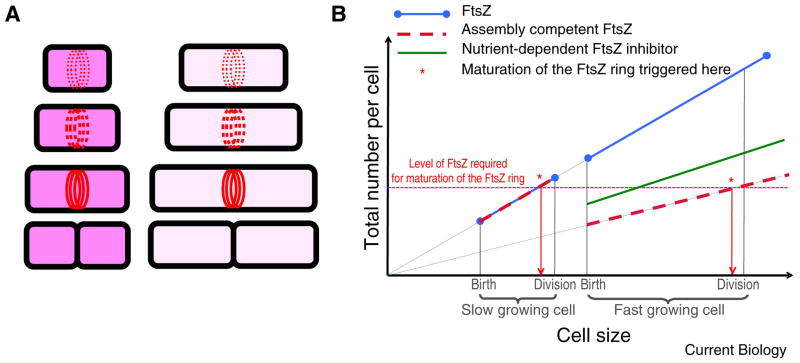
Like eukaryotes, bacteria must coordinate division with growth to ensure cells are the appropriate size for a given environmental condition or developmental fate. As single-celled organisms, nutrient availability is one of the strongest influences on bacterial cell size. Classic physiological experiments conducted over four decades ago first demonstrated that cell size is directly correlated with nutrient source and growth rate in the Gram-negative bacterium Salmonella typhimurium. This observation subsequently served as the basis for studies revealing a role for cell size in cell cycle progression in a closely related organism, Escherichia coli. More recently, the development of powerful genetic, molecular, and imaging tools has allowed us to identify and characterize the nutrient-dependent pathway responsible for coordinating cell division and cell size with growth rate in the Gram-positive model organism Bacillus subtilis. Here, we discuss the role of cell size in bacterial growth and development and propose a broadly applicable model for cell size control in this important and highly divergent domain of life.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Grover NB, Woldringh CL. Dimensional regulation of cell-cycle events in Escherichia coli during steady-state growth. Microbiology. 2001;147:171–181. - PubMed
-
- Angert ER, Clements KD, Pace NR. The largest bacterium. Nature. 1993;362:239–241. - PubMed
-
- Schulz HN, Jorgensen BB. Big bacteria. Annu Rev Microbiol. 2001;55:105–137. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
