

Cellular and physiological mechanisms underlying blood flow regulation in the retina and choroid in health and disease
- PMID: 22580107
- PMCID: PMC3418965
- DOI: 10.1016/j.preteyeres.2012.04.004
Cellular and physiological mechanisms underlying blood flow regulation in the retina and choroid in health and disease
Abstract
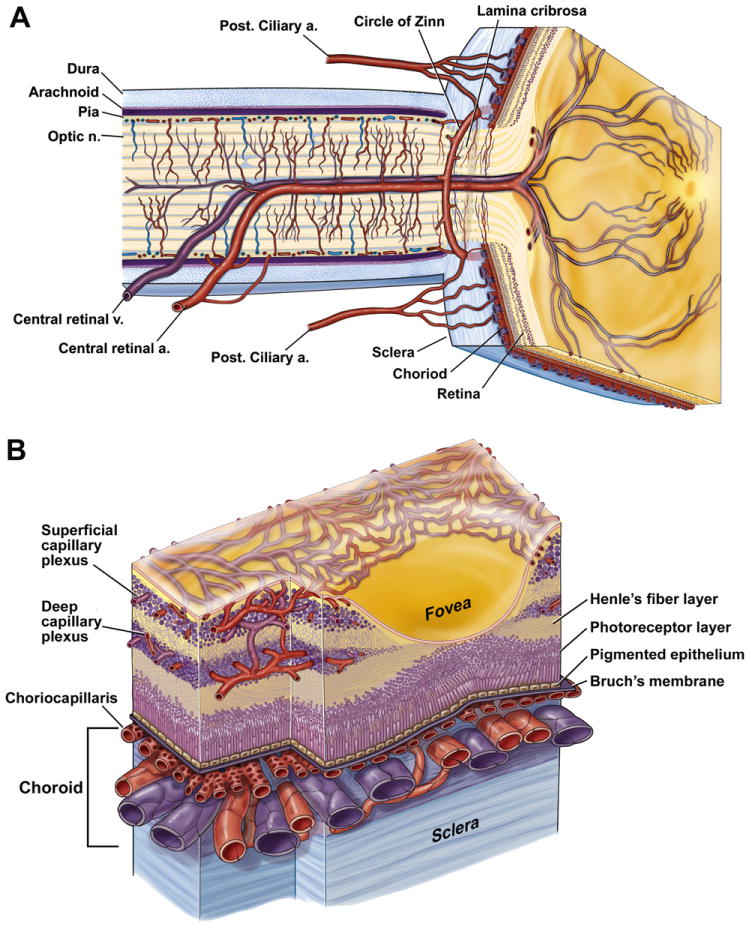
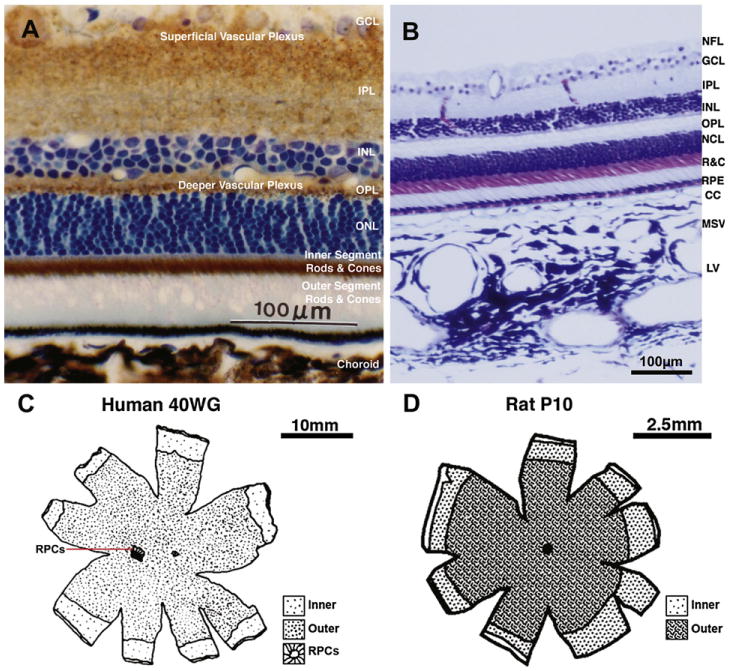
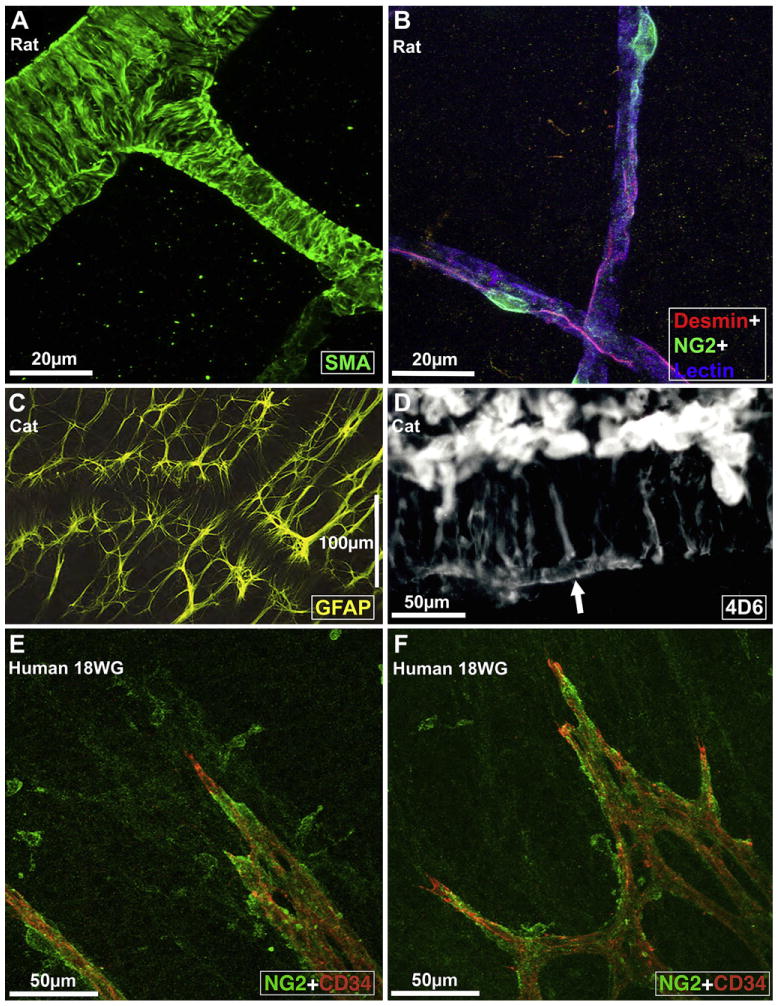
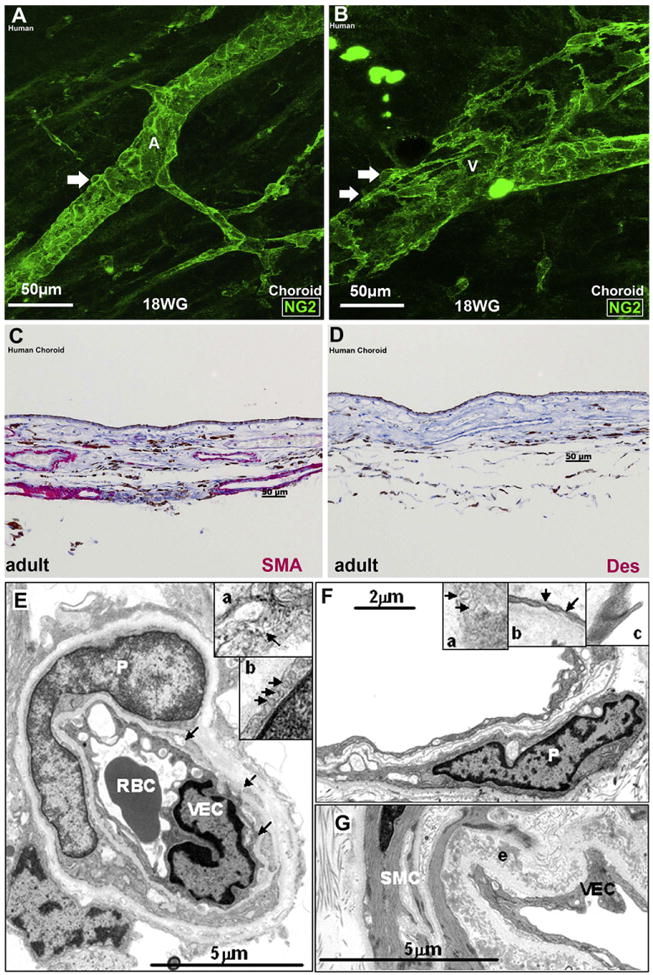
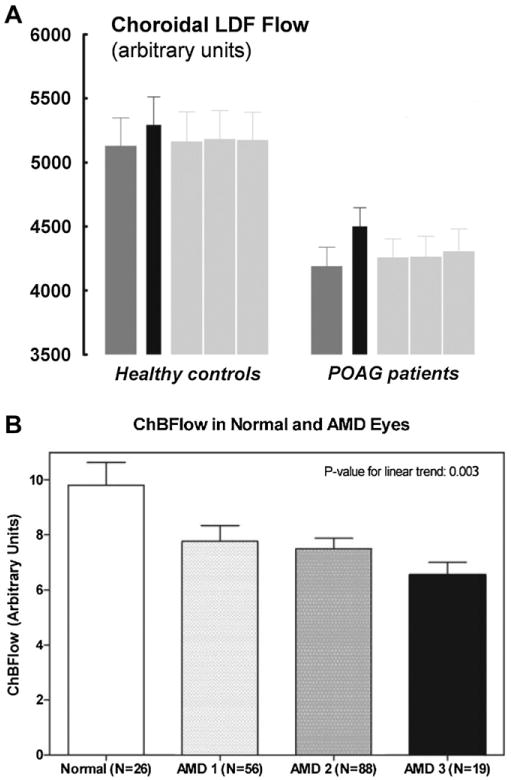
We review the cellular and physiological mechanisms responsible for the regulation of blood flow in the retina and choroid in health and disease. Due to the intrinsic light sensitivity of the retina and the direct visual accessibility of fundus blood vessels, the eye offers unique opportunities for the non-invasive investigation of mechanisms of blood flow regulation. The ability of the retinal vasculature to regulate its blood flow is contrasted with the far more restricted ability of the choroidal circulation to regulate its blood flow by virtue of the absence of glial cells, the markedly reduced pericyte ensheathment of the choroidal vasculature, and the lack of intermediate filaments in choroidal pericytes. We review the cellular and molecular components of the neurovascular unit in the retina and choroid, techniques for monitoring retinal and choroidal blood flow, responses of the retinal and choroidal circulation to light stimulation, the role of capillaries, astrocytes and pericytes in regulating blood flow, putative signaling mechanisms mediating neurovascular coupling in the retina, and changes that occur in the retinal and choroidal circulation during diabetic retinopathy, age-related macular degeneration, glaucoma, and Alzheimer's disease. We close by discussing issues that remain to be explored.
Crown Copyright © 2012. Published by Elsevier Ltd. All rights reserved.
Figures
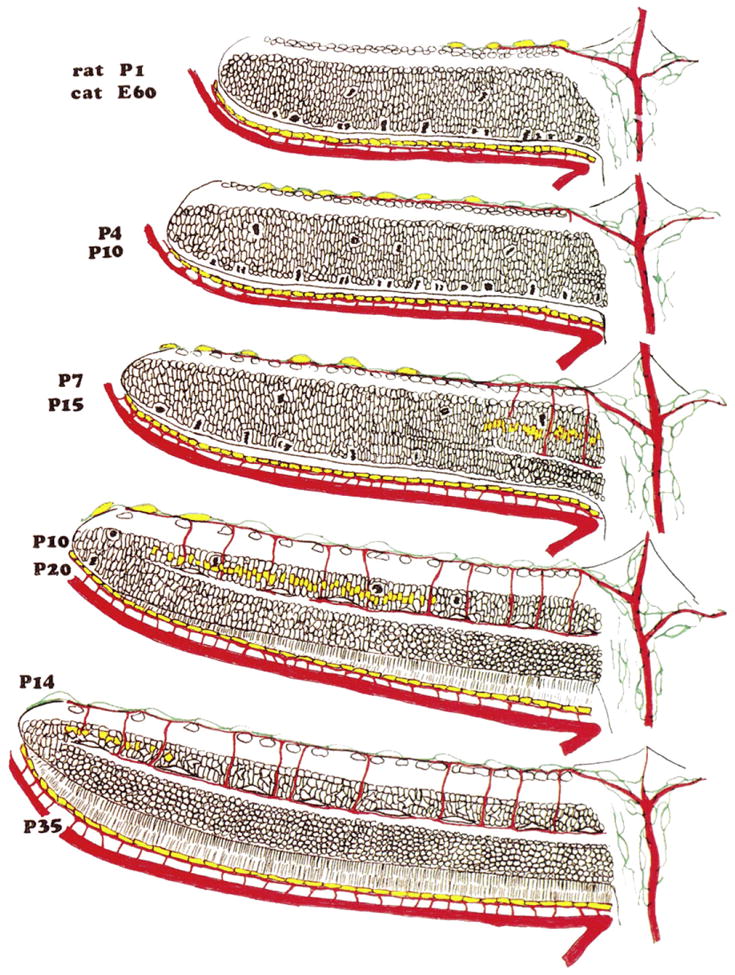
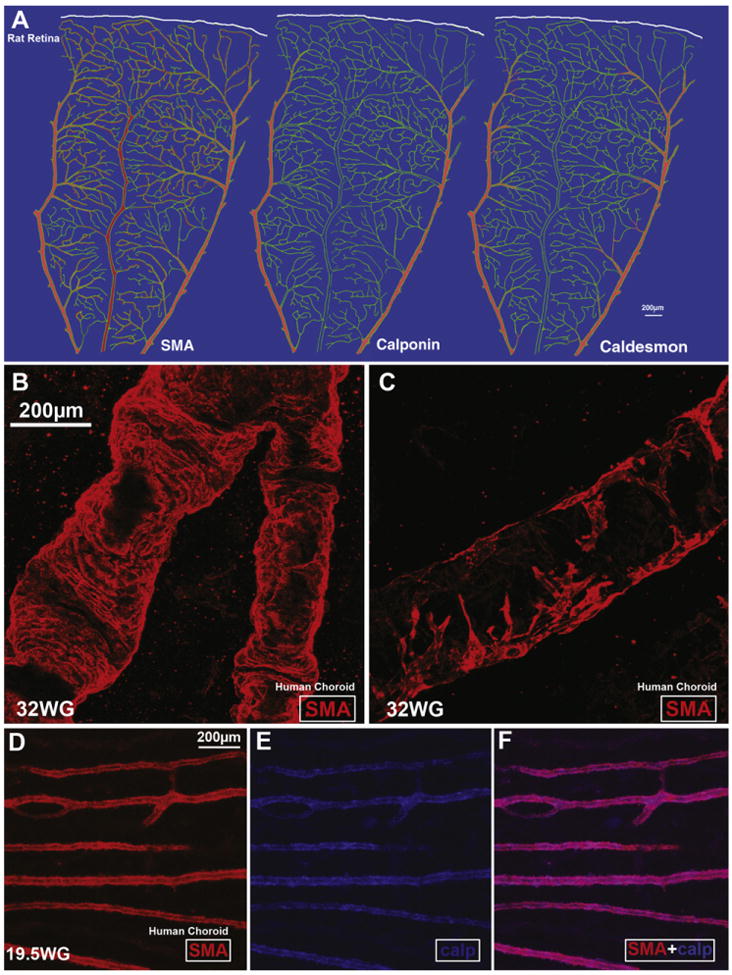
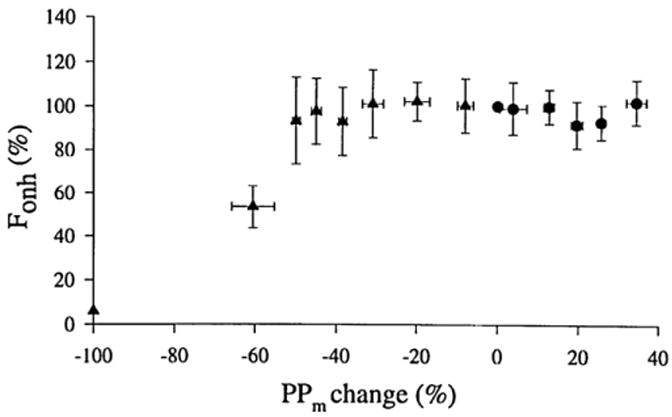
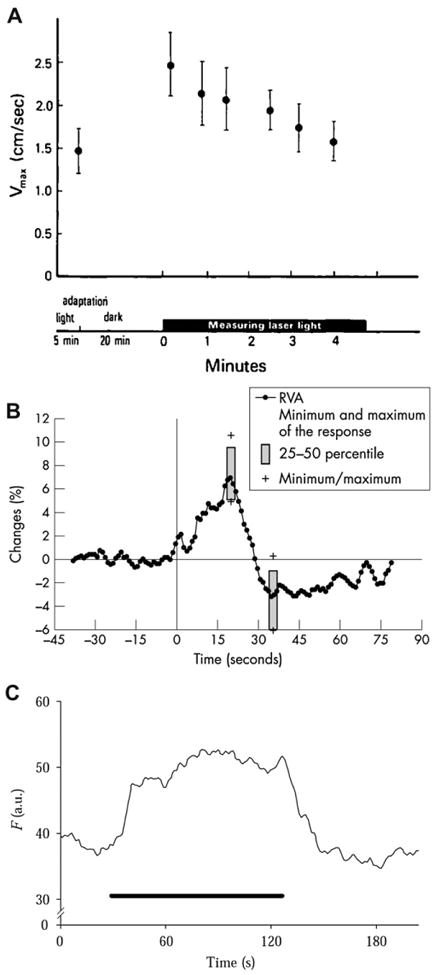
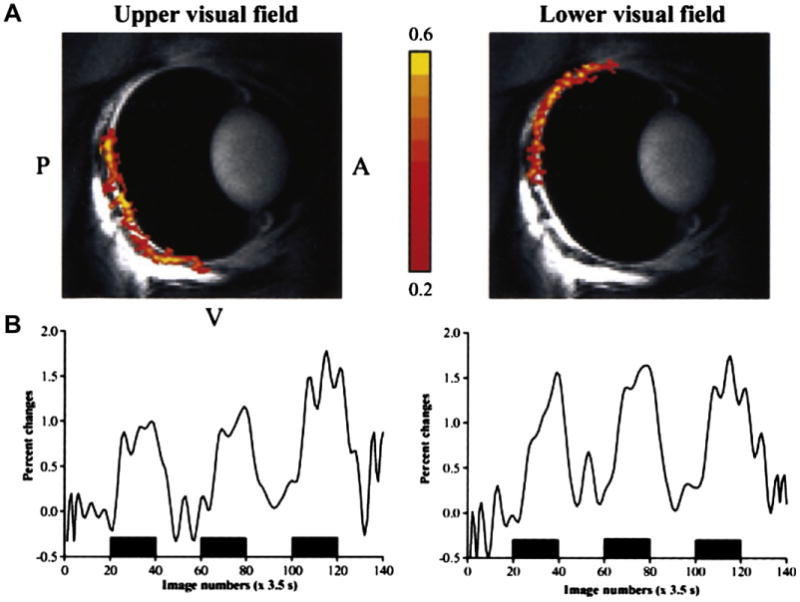
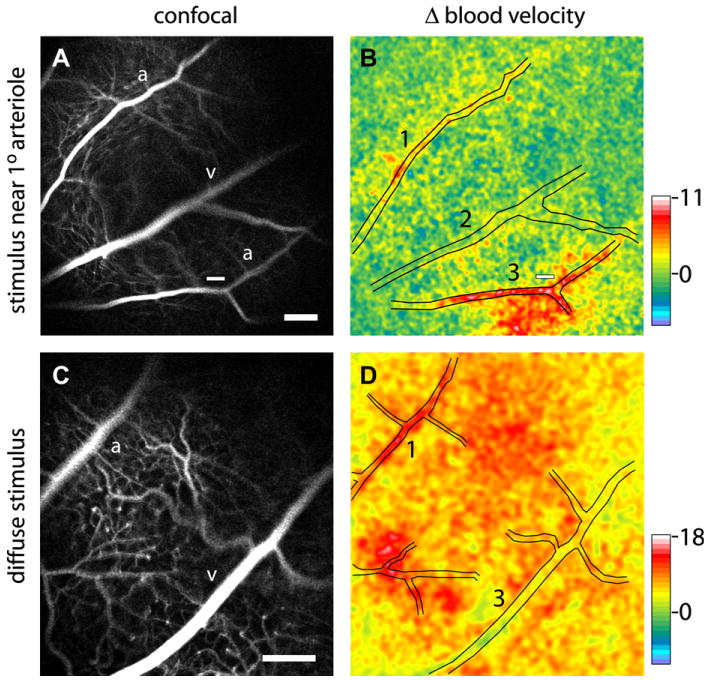

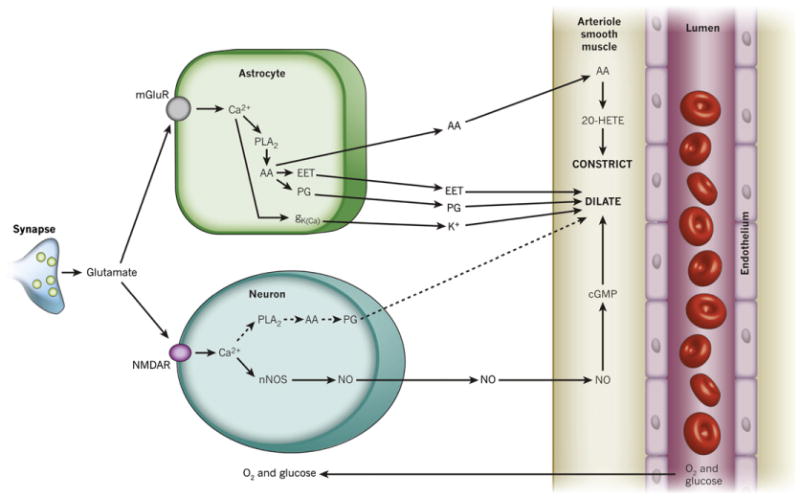
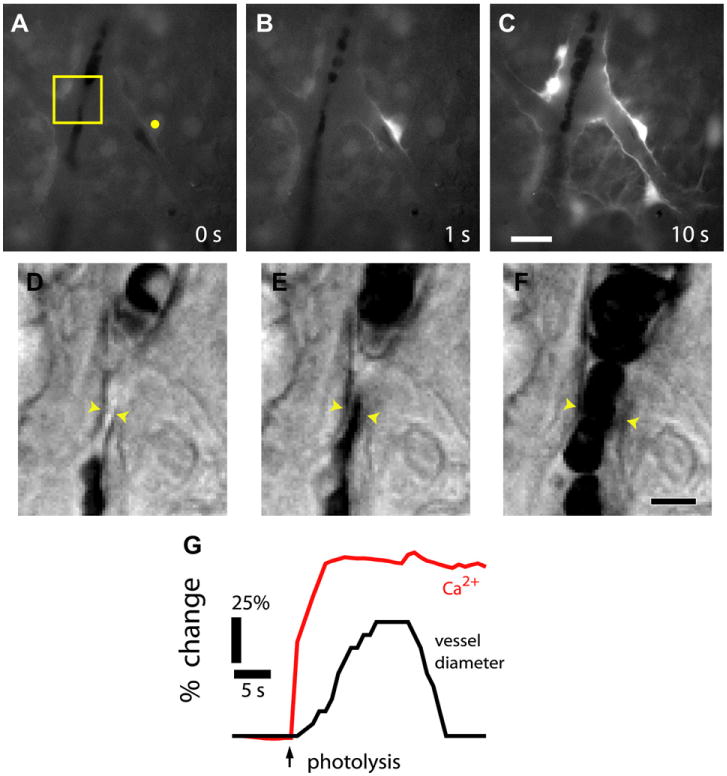
References
-
- Akata T. Cellular and molecular mechanisms regulating vascular tone. Part 2: regulatory mechanisms modulating Ca2+ mobilization and/or myofilament Ca2+ sensitivity in vascular smooth muscle cells. J Anesth. 2007;21:232–242. - PubMed
-
- Alder VA, Ben-Nun J, Cringle SJ. PO2 profiles and oxygen consumption in cat retina with an occluded retinal circulation. Invest Ophthalmol Vis Sci. 1990;31:1029–1034. - PubMed
-
- Alm A, Bill A. The oxygen supply to the retina. I. Effects of changes in intraocular and arterial blood pressures, and in arterial PO2 and PCO2 on the oxygen tension in the vitreous body of the cat. Acta Physiol Scand. 1972a;84:261–274. - PubMed
-
- Alm A, Bill A. The oxygen supply to the retina. II. Effects of high intraocular pressure and of increased arterial carbon dioxide tension on uveal and retinal blood flow in cats. A study with radioactively labelled microspheres including flow determinations in brain and some other tissues. Acta Physiol Scand. 1972b;84:306–319. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
