

ApoB-containing lipoproteins regulate angiogenesis by modulating expression of VEGF receptor 1
- PMID: 22581286
- PMCID: PMC3959651
- DOI: 10.1038/nm.2759
ApoB-containing lipoproteins regulate angiogenesis by modulating expression of VEGF receptor 1
Abstract
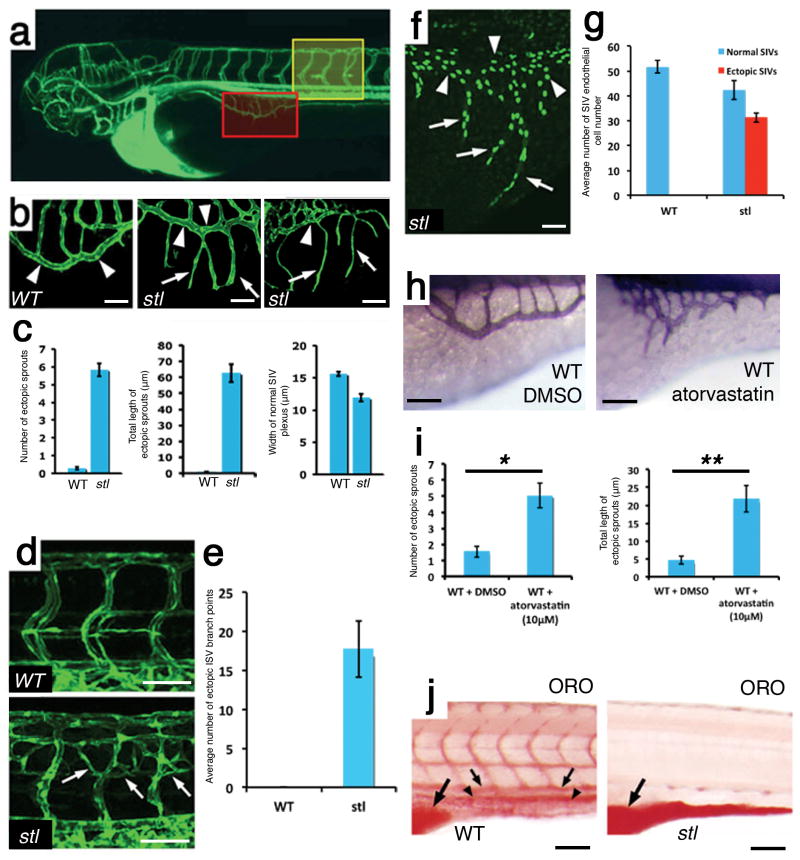
Despite the clear major contribution of hyperlipidemia to the prevalence of cardiovascular disease in the developed world, the direct effects of lipoproteins on endothelial cells have remained obscure and are under debate. Here we report a previously uncharacterized mechanism of vessel growth modulation by lipoprotein availability. Using a genetic screen for vascular defects in zebrafish, we initially identified a mutation, stalactite (stl), in the gene encoding microsomal triglyceride transfer protein (mtp), which is involved in the biosynthesis of apolipoprotein B (ApoB)-containing lipoproteins. By manipulating lipoprotein concentrations in zebrafish, we found that ApoB negatively regulates angiogenesis and that it is the ApoB protein particle, rather than lipid moieties within ApoB-containing lipoproteins, that is primarily responsible for this effect. Mechanistically, we identified downregulation of vascular endothelial growth factor receptor 1 (VEGFR1), which acts as a decoy receptor for VEGF, as a key mediator of the endothelial response to lipoproteins, and we observed VEGFR1 downregulation in hyperlipidemic mice. These findings may open new avenues for the treatment of lipoprotein-related vascular disorders.
Conflict of interest statement
The authors declare no competing financial interests.
Figures



References
-
- Yaniv K, et al. Live imaging of lymphatic development in the zebrafish. Nat Med. 2006;12:711–716. - PubMed
-
- Lawson ND, Weinstein BM. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev Biol. 2002;248:307–318. - PubMed
-
- Hussain MM, et al. Microsomal triglyceride transfer protein in plasma and cellular lipid metabolism. Curr Opin Lipidol. 2008;19:277–284. - PubMed
-
- Marza E, et al. Developmental expression and nutritional regulation of a zebrafish gene homologous to mammalian microsomal triglyceride transfer protein large subunit. Dev Dyn. 2005;232:506–518. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
