

Changes in descending motor pathway connectivity after corticospinal tract lesion in macaque monkey
- PMID: 22581799
- PMCID: PMC3381720
- DOI: 10.1093/brain/aws115
Changes in descending motor pathway connectivity after corticospinal tract lesion in macaque monkey
Abstract
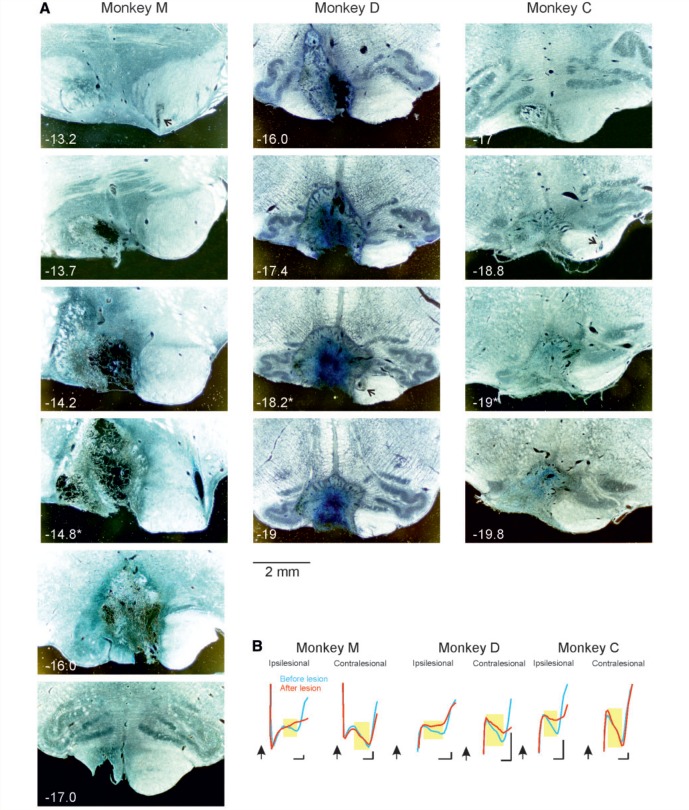
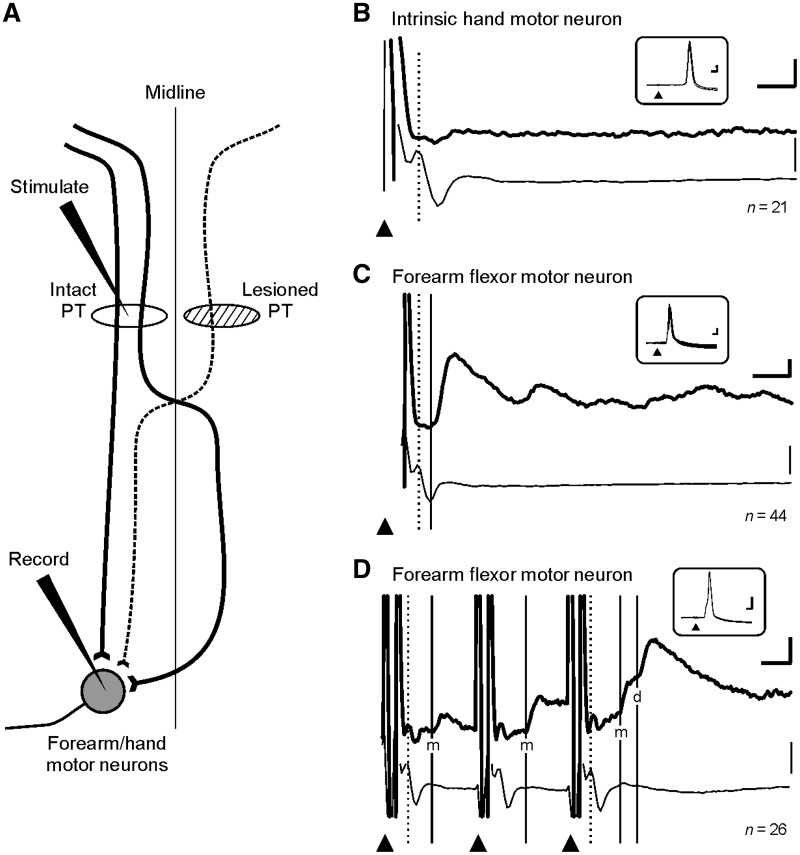
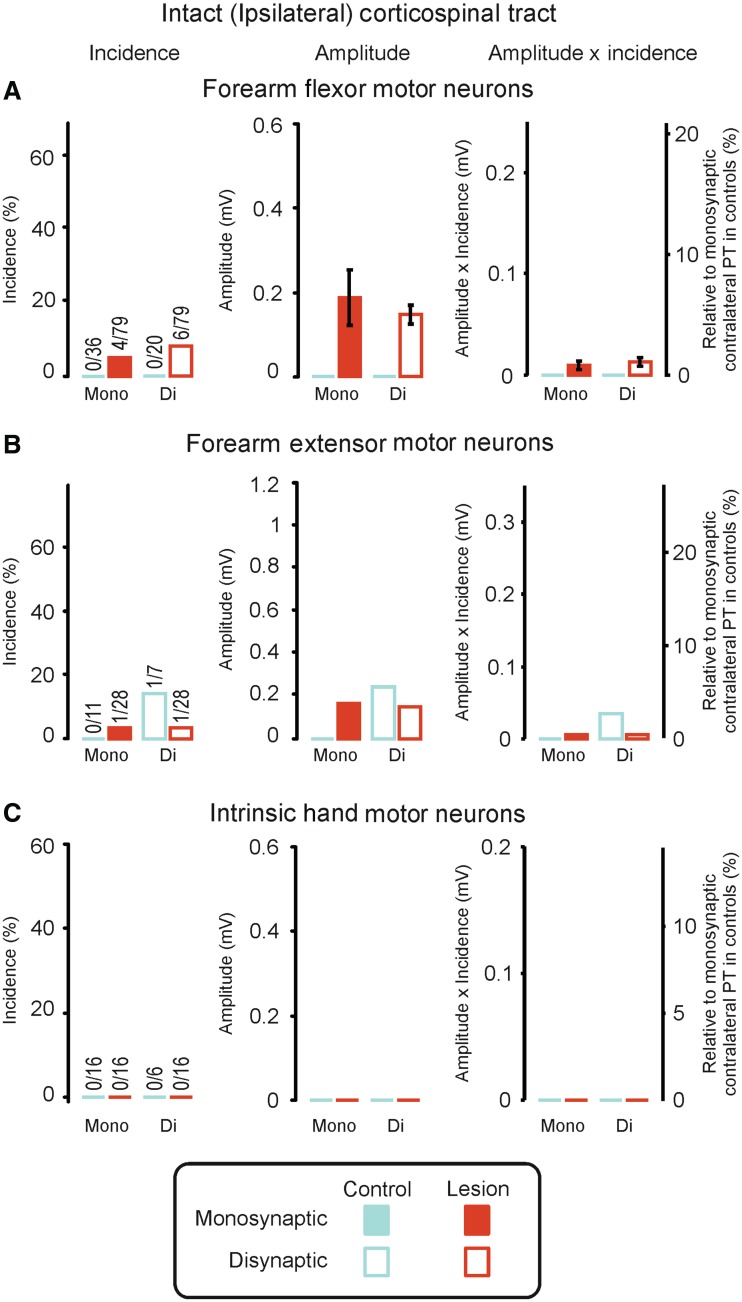
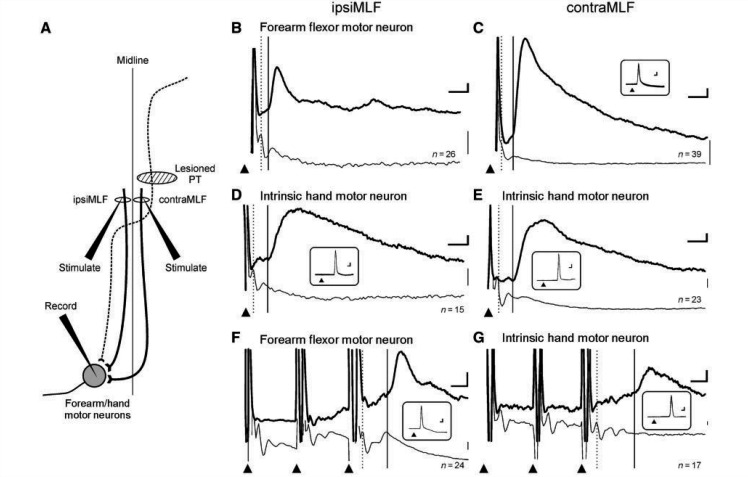
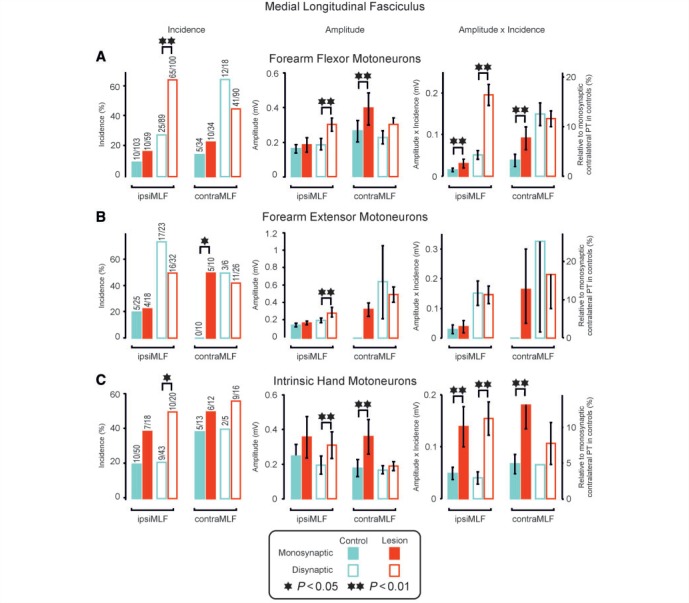
Damage to the corticospinal tract is a leading cause of motor disability, for example in stroke or spinal cord injury. Some function usually recovers, but whether plasticity of undamaged ipsilaterally descending corticospinal axons and/or brainstem pathways such as the reticulospinal tract contributes to recovery is unknown. Here, we examined the connectivity in these pathways to motor neurons after recovery from corticospinal lesions. Extensive unilateral lesions of the medullary corticospinal fibres in the pyramidal tract were made in three adult macaque monkeys. After an initial contralateral flaccid paralysis, motor function rapidly recovered, after which all animals were capable of climbing and supporting their weight by gripping the cage bars with the contralesional hand. In one animal where experimental testing was carried out, there was (as expected) no recovery of fine independent finger movements. Around 6 months post-lesion, intracellular recordings were made from 167 motor neurons innervating hand and forearm muscles. Synaptic responses evoked by stimulating the unlesioned ipsilateral pyramidal tract and the medial longitudinal fasciculus were recorded and compared with control responses in 207 motor neurons from six unlesioned animals. Input from the ipsilateral pyramidal tract was rare and weak in both lesioned and control animals, suggesting a limited role for this pathway in functional recovery. In contrast, mono- and disynaptic excitatory post-synaptic potentials elicited from the medial longitudinal fasciculus significantly increased in average size after recovery, but only in motor neurons innervating forearm flexor and intrinsic hand muscles, not in forearm extensor motor neurons. We conclude that reticulospinal systems sub-serve some of the functional recovery after corticospinal lesions. The imbalanced strengthening of connections to flexor, but not extensor, motor neurons mirrors the extensor weakness and flexor spasm which in neurological experience is a common limitation to recovery in stroke survivors.
Figures
Similar articles
-
Does a C3-C4 propriospinal system transmit corticospinal excitation in the primate? An investigation in the macaque monkey.J Physiol. 1998 Aug 15;511 ( Pt 1)(Pt 1):191-212. doi: 10.1111/j.1469-7793.1998.191bi.x. J Physiol. 1998. PMID: 9679174 Free PMC article.
-
Direct and indirect connections with upper limb motoneurons from the primate reticulospinal tract.J Neurosci. 2009 Apr 15;29(15):4993-9. doi: 10.1523/JNEUROSCI.3720-08.2009. J Neurosci. 2009. PMID: 19369568 Free PMC article.
-
Uncrossed actions of feline corticospinal tract neurones on hindlimb motoneurones evoked via ipsilaterally descending pathways.J Physiol. 2007 Apr 1;580(Pt 1):119-32. doi: 10.1113/jphysiol.2006.122721. Epub 2007 Jan 25. J Physiol. 2007. PMID: 17255171 Free PMC article.
-
How can corticospinal tract neurons contribute to ipsilateral movements? A question with implications for recovery of motor functions.Neuroscientist. 2006 Feb;12(1):67-79. doi: 10.1177/1073858405283392. Neuroscientist. 2006. PMID: 16394194 Free PMC article. Review.
-
[The corticospinal tract and its role in motor control].Brain Nerve. 2012 Nov;64(11):1331-9. Brain Nerve. 2012. PMID: 23131745 Review. Japanese.
Cited by
-
What are the Best Animal Models for Testing Early Intervention in Cerebral Palsy?Front Neurol. 2014 Dec 4;5:258. doi: 10.3389/fneur.2014.00258. eCollection 2014. Front Neurol. 2014. PMID: 25538677 Free PMC article. Review.
-
The Gigantocellular Reticular Nucleus Plays a Significant Role in Locomotor Recovery after Incomplete Spinal Cord Injury.J Neurosci. 2020 Oct 21;40(43):8292-8305. doi: 10.1523/JNEUROSCI.0474-20.2020. Epub 2020 Sep 25. J Neurosci. 2020. PMID: 32978289 Free PMC article.
-
Decoding upper limb residual muscle activity in severe chronic stroke.Ann Clin Transl Neurol. 2015 Jan;2(1):1-11. doi: 10.1002/acn3.122. Epub 2014 Dec 9. Ann Clin Transl Neurol. 2015. PMID: 25642429 Free PMC article.
-
Variable Interhemispheric Asymmetry in Layer V of the Supplementary Motor Area following Cervical Hemisection in Adult Macaque Monkeys.eNeuro. 2020 Oct 9;7(5):ENEURO.0280-20.2020. doi: 10.1523/ENEURO.0280-20.2020. Print 2020 Sep/Oct. eNeuro. 2020. PMID: 32917794 Free PMC article.
-
Neural Substrates of Motor Recovery in Severely Impaired Stroke Patients With Hand Paralysis.Neurorehabil Neural Repair. 2016 May;30(4):328-38. doi: 10.1177/1545968315594886. Epub 2015 Jul 10. Neurorehabil Neural Repair. 2016. PMID: 26163204 Free PMC article.
References
-
- Alstermark B, Isa T, Ohki Y, Saito Y. Disynaptic pyramidal excitation in forelimb motoneurons mediated via C(3)-C(4) propriospinal neurons in the Macaca fuscata. J Neurophysiol. 1999;82:3580–5. - PubMed
-
- Bareyre FM, Kerschensteiner M, Raineteau O, Mettenleiter TC, Weinmann O, Schwab ME. The injured spinal cord spontaneously forms a new intraspinal circuit in adult rats. Nat Neurosci. 2004;7:269–77. - PubMed
-
- Belhaj-Saif A, Cheney PD. Plasticity in the distribution of the red nucleus output to forearm muscles after unilateral lesions of the pyramidal tract. J Neurophysiol. 2000;83:3147–53. - PubMed
-
- Benecke R, Meyer BU, Freund HJ. Reorganisation of descending motor pathways in patients after hemispherectomy and severe hemispheric lesions demonstrated by magnetic brain stimulation. Exp Brain Res. 1991;83:419–26. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
