

Regulatory effects of programmed cell death 4 (PDCD4) protein in interferon (IFN)-stimulated gene expression and generation of type I IFN responses
- PMID: 22586265
- PMCID: PMC3416205
- DOI: 10.1128/MCB.00310-12
Regulatory effects of programmed cell death 4 (PDCD4) protein in interferon (IFN)-stimulated gene expression and generation of type I IFN responses
Erratum in
- Mol Cell Biol. 2012 Sep;32(17):3608
Abstract
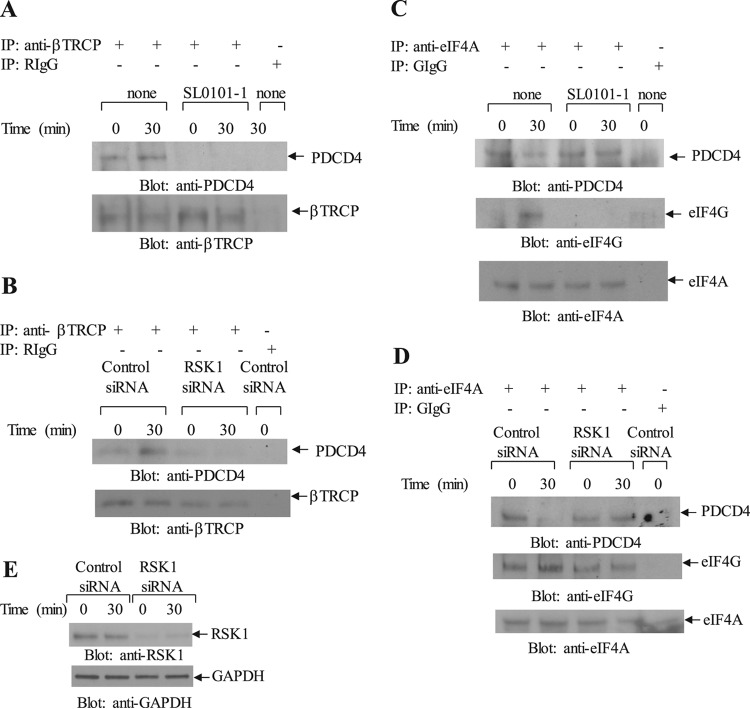
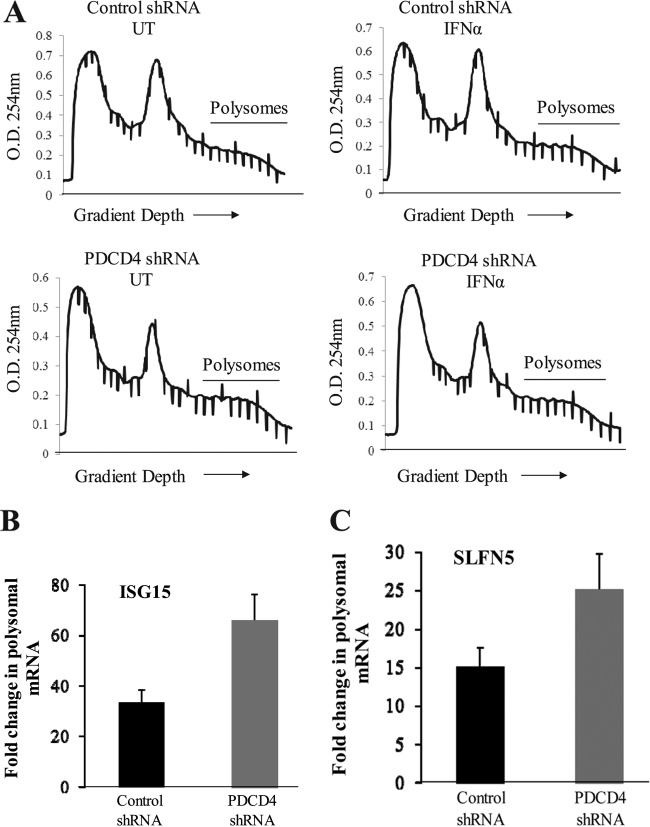
The precise mechanisms by which the activation of interferon (IFN) receptors (IFNRs) ultimately controls mRNA translation of specific target genes to induce IFN-dependent biological responses remain ill defined. We provide evidence that IFN-α induces phosphorylation of programmed cell death 4 (PDCD4) protein on Ser67. This IFN-α-dependent phosphorylation is mediated by either the p70 S6 kinase (S6K) or the p90 ribosomal protein S6K (RSK) in a cell-type-specific manner. IFN-dependent phosphorylation of PDCD4 results in downregulation of PDCD4 protein levels as the phosphorylated form of PDCD4 interacts with the ubiquitin ligase β-TRCP (β-transducin repeat-containing protein) and undergoes degradation. This process facilitates IFN-induced eukaryotic translation initiation factor 4A (eIF4A) activity and binding to translation initiation factor eIF4G to promote mRNA translation. Our data establish that PDCD4 degradation ultimately facilitates expression of several ISG protein products that play important roles in the generation of IFN responses, including IFN-stimulated gene 15 (ISG15), p21(WAF1/CIP1), and Schlafen 5 (SLFN5). Moreover, engagement of the RSK/PDCD4 pathway by the type I IFNR is required for the suppressive effects of IFN-α on normal CD34(+) hematopoietic precursors and for antileukemic effects in vitro. Altogether, these findings provide evidence for a unique function of PDCD4 in the type I IFN system and indicate a key regulatory role for this protein in mRNA translation of ISGs and control of IFN responses.
Figures









Similar articles
-
Interferon-dependent engagement of eukaryotic initiation factor 4B via S6 kinase (S6K)- and ribosomal protein S6K-mediated signals.Mol Cell Biol. 2009 May;29(10):2865-75. doi: 10.1128/MCB.01537-08. Epub 2009 Mar 16. Mol Cell Biol. 2009. PMID: 19289497 Free PMC article.
-
Suppression of programmed cell death 4 (PDCD4) protein expression by BCR-ABL-regulated engagement of the mTOR/p70 S6 kinase pathway.J Biol Chem. 2008 Mar 28;283(13):8601-10. doi: 10.1074/jbc.M707934200. Epub 2008 Jan 26. J Biol Chem. 2008. PMID: 18223253 Free PMC article.
-
S6K1- and betaTRCP-mediated degradation of PDCD4 promotes protein translation and cell growth.Science. 2006 Oct 20;314(5798):467-71. doi: 10.1126/science.1130276. Science. 2006. PMID: 17053147
-
Programmed cell death protein 4 (pdcd4): a novel target for antineoplastic therapy?Biol Cell. 2003 Nov;95(8):515-9. doi: 10.1016/j.biolcel.2003.09.003. Biol Cell. 2003. PMID: 14630388 Review.
-
The tumour suppressor Pdcd4: recent advances in the elucidation of function and regulation.Biol Cell. 2009 Jun;101(6):309-17. doi: 10.1042/BC20080191. Biol Cell. 2009. PMID: 19356152 Review.
Cited by
-
Host translation at the nexus of infection and immunity.Cell Host Microbe. 2012 Oct 18;12(4):470-83. doi: 10.1016/j.chom.2012.09.006. Cell Host Microbe. 2012. PMID: 23084916 Free PMC article. Review.
-
Regulation of interferon-dependent mRNA translation of target genes.J Interferon Cytokine Res. 2014 Apr;34(4):289-96. doi: 10.1089/jir.2013.0148. Epub 2014 Feb 21. J Interferon Cytokine Res. 2014. PMID: 24559173 Free PMC article. Review.
-
Interferon γ (IFNγ) Signaling via Mechanistic Target of Rapamycin Complex 2 (mTORC2) and Regulatory Effects in the Generation of Type II Interferon Biological Responses.J Biol Chem. 2016 Jan 29;291(5):2389-96. doi: 10.1074/jbc.M115.664995. Epub 2015 Dec 8. J Biol Chem. 2016. PMID: 26645692 Free PMC article.
-
MicroRNA-21: A Positive Regulator for Optimal Production of Type I and Type III Interferon by Plasmacytoid Dendritic Cells.Front Immunol. 2017 Aug 21;8:947. doi: 10.3389/fimmu.2017.00947. eCollection 2017. Front Immunol. 2017. PMID: 28871250 Free PMC article.
-
Identification of Key Deregulated RNA-Binding Proteins in Pancreatic Cancer by Meta-Analysis and Prediction of Their Role as Modulators of Oncogenesis.Front Cell Dev Biol. 2021 Nov 29;9:713852. doi: 10.3389/fcell.2021.713852. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34912796 Free PMC article.
References
-
- Bitomsky N, Wethkamp N, Marikkannu R, Klempnauer KH. 2008. siRNA-mediated knockdown of Pdcd4 expression causes up regulation of p21Waf1/Cip1 expression. Oncogene 27:4820–4829 - PubMed
-
- Bohm M, et al. 2003. The transformation suppressor protein Pdcd4 shuttles between nucleus and cytoplasm and binds RNA. Oncogene 22:4905–4910 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous