

A roundabout way to cancer
- PMID: 22588058
- PMCID: PMC4121377
- DOI: 10.1016/B978-0-12-386503-8.00005-3
A roundabout way to cancer
Abstract
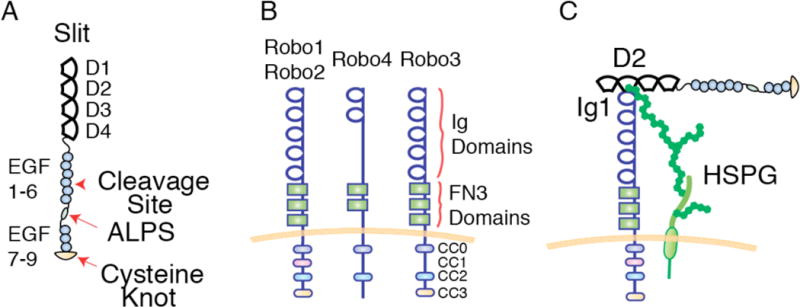
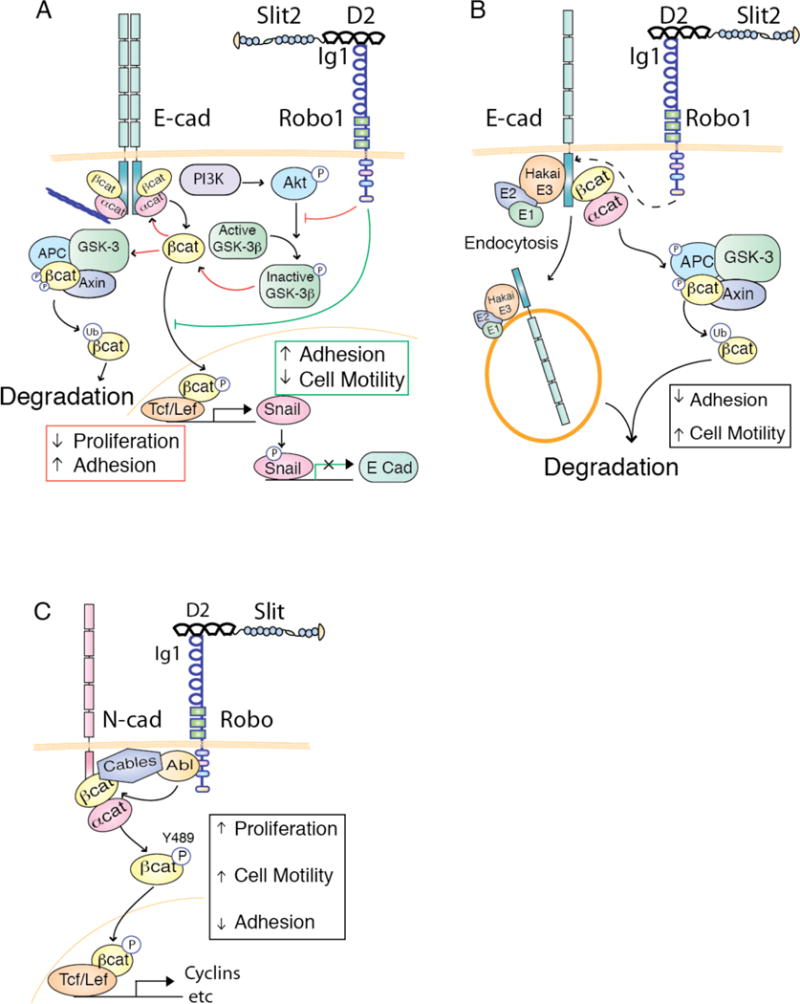
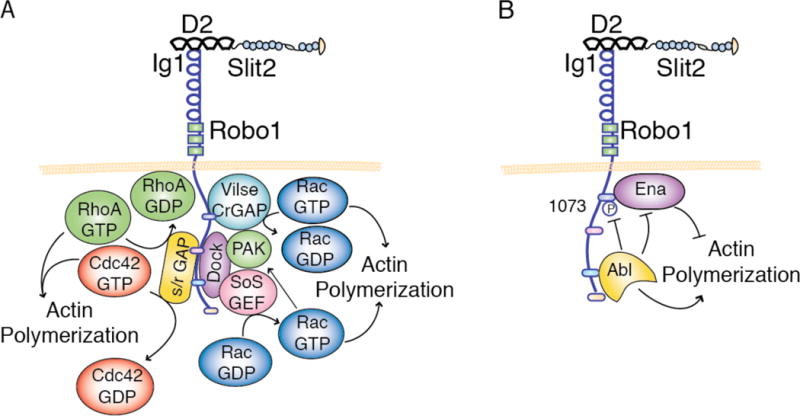
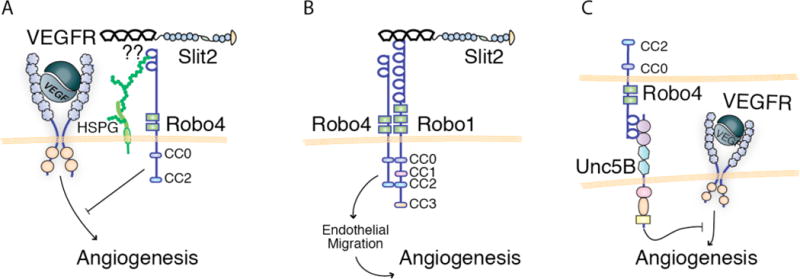
The Slit family of secreted proteins and their transmembrane receptor, Robo, were originally identified in the nervous system where they function as axon guidance cues and branching factors during development. Since their discovery, a great number of additional roles have been attributed to Slit/Robo signaling, including regulating the critical processes of cell proliferation and cell motility in a variety of cell and tissue types. These processes are often deregulated during cancer progression, allowing tumor cells to bypass safeguarding mechanisms in the cell and the environment in order to grow and escape to new tissues. In the past decade, it has been shown that the expression of Slit and Robo is altered in a wide variety of cancer types, identifying them as potential therapeutic targets. Further, studies have demonstrated dual roles for Slits and Robos in cancer, acting as both oncogenes and tumor suppressors. This bifunctionality is also observed in their roles as axon guidance cues in the developing nervous system, where they both attract and repel neuronal migration. The fact that this signaling axis can have opposite functions depending on the cellular circumstance make its actions challenging to define. Here, we summarize our current understanding of the dual roles that Slit/Robo signaling play in development, epithelial tumor progression, and tumor angiogenesis.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Alajez NM, Lenarduzzi M, Ito E, Hui AB, Shi W, Bruce J, Yue S, Huang SH, Xu W, Waldron J, O’Sullivan B, Liu FF. MiR-218 suppresses nasopharyngeal cancer progression through downregulation of survivin and the SLIT2-ROBO1 pathway. Cancer Res. 2011;71:2381–2391. - PubMed
-
- Angeloni D, ter Elst A, Wei MH, van der Veen AY, Braga EA, Klimov EA, Timmer T, Korobeinikova L, Lerman MI, Buys CH. Analysis of a new homozygous deletion in the tumor suppressor region at 3p12.3 reveals two novel intronic noncoding RNA genes. Genes Chromosomes Cancer. 2006;45:676–691. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
