

Suppression of heat shock protein 27 induces long-term dormancy in human breast cancer
- PMID: 22589302
- PMCID: PMC3365195
- DOI: 10.1073/pnas.1017909109
Suppression of heat shock protein 27 induces long-term dormancy in human breast cancer
Abstract
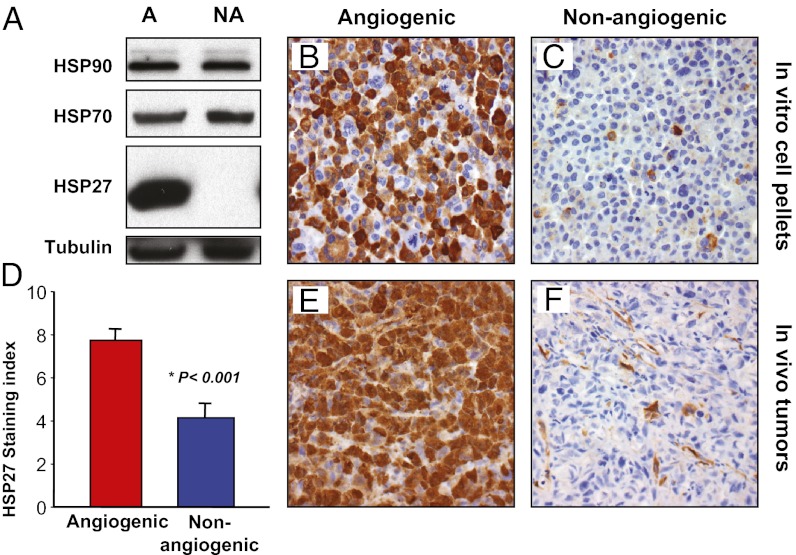
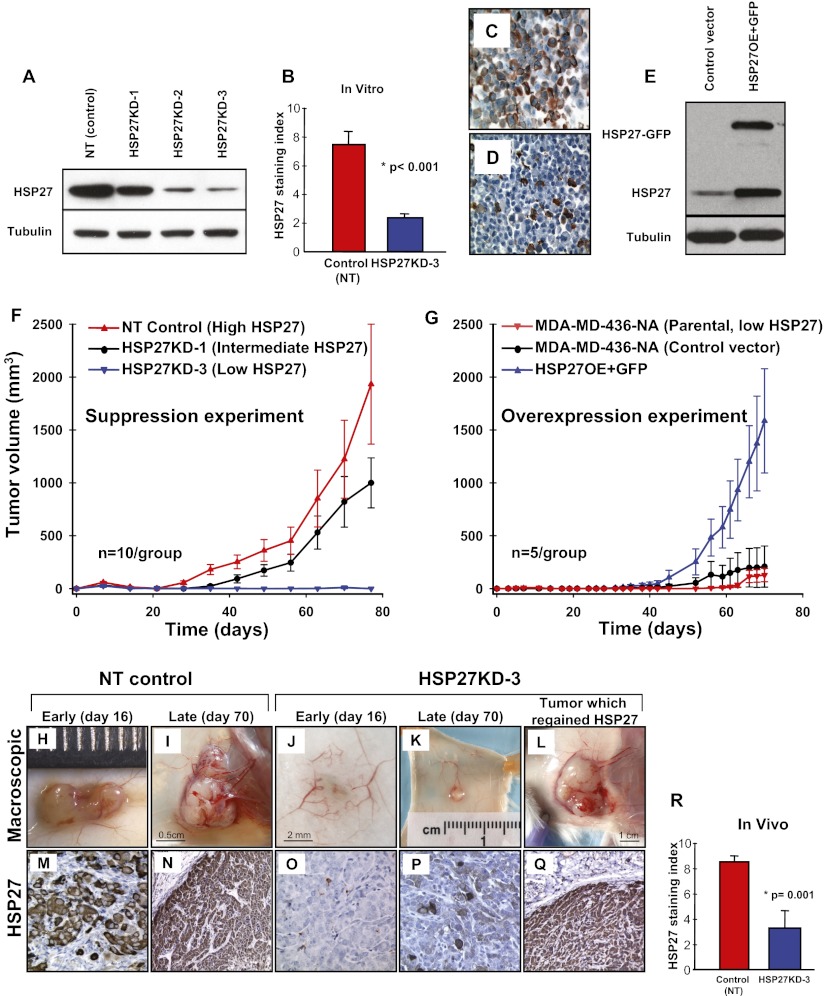
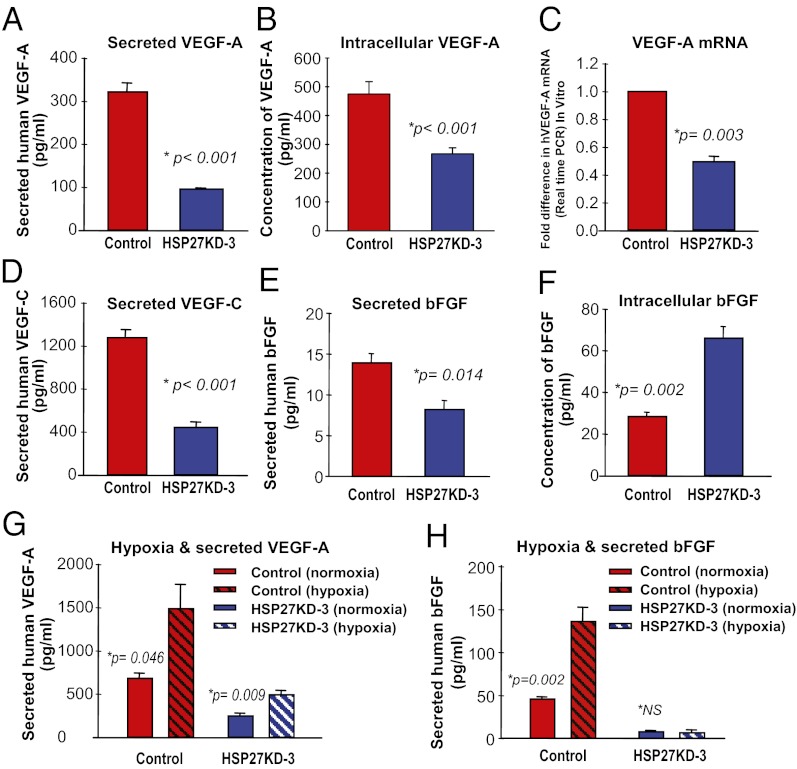
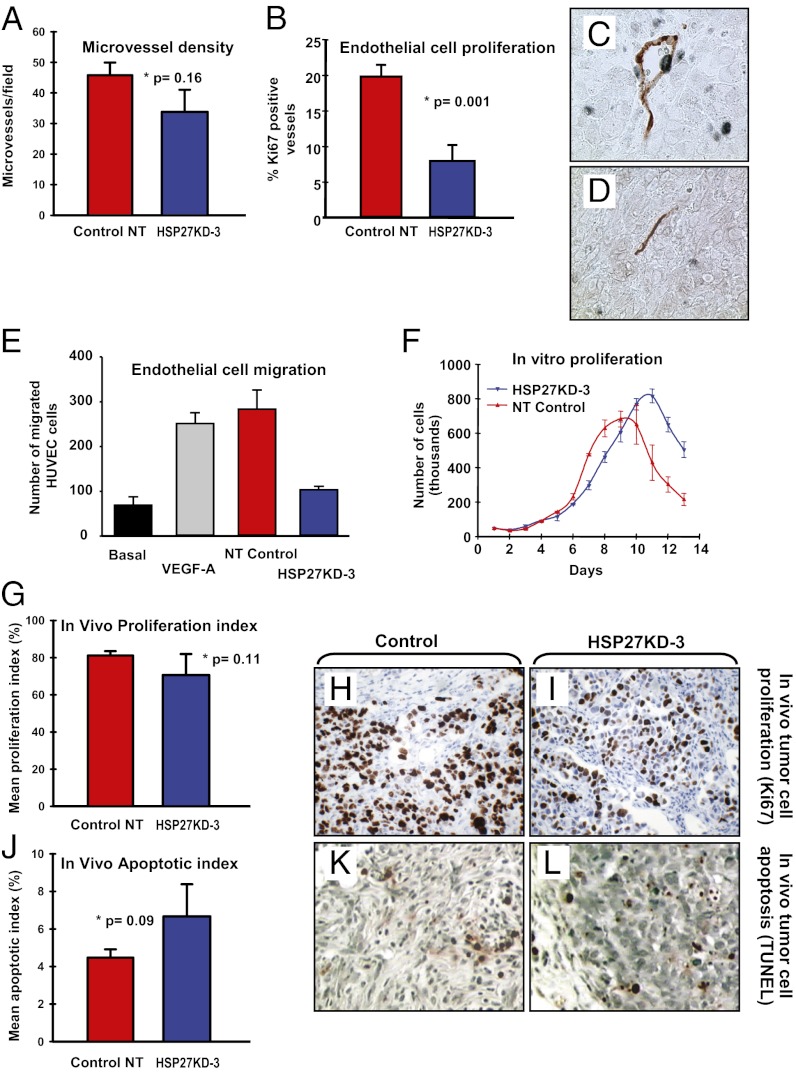
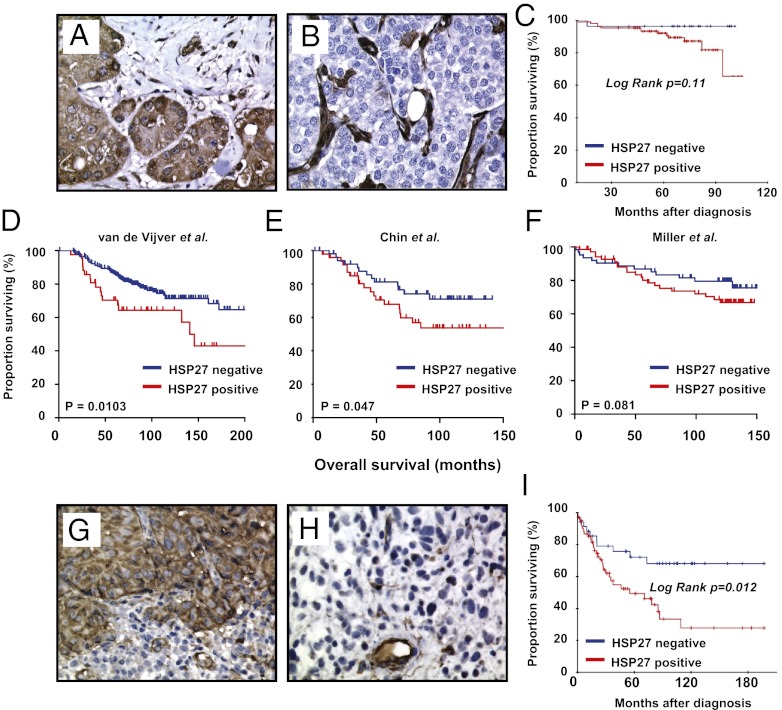
The mechanisms underlying tumor dormancy have been elusive and not well characterized. We recently published an experimental model for the study of human tumor dormancy and the role of angiogenesis, and reported that the angiogenic switch was preceded by a local increase in VEGF-A and basic fibroblast growth factor. In this breast cancer xenograft model (MDA-MB-436 cells), analysis of differentially expressed genes revealed that heat shock protein 27 (HSP27) was significantly up-regulated in angiogenic cells compared with nonangiogenic cells. The effect of HSP27 down-regulation was further evaluated in cell lines, mouse models, and clinical datasets of human patients with breast cancer and melanoma. Stable down-regulation of HSP27 in angiogenic tumor cells was followed by long-term tumor dormancy in vivo. Strikingly, only 4 of 30 HSP27 knockdown xenograft tumors initiated rapid growth after day 70, in correlation with a regain of HSP27 protein expression. Significantly, no tumors escaped from dormancy without HSP27 expression. Down-regulation of HSP27 was associated with reduced endothelial cell proliferation and decreased secretion of VEGF-A, VEGF-C, and basic fibroblast growth factor. Conversely, overexpression of HSP27 in nonangiogenic cells resulted in expansive tumor growth in vivo. By clinical validation, strong HSP27 protein expression was associated with markers of aggressive tumors and decreased survival in patients with breast cancer and melanoma. An HSP27-associated gene expression signature was related to molecular subgroups and survival in breast cancer. Our findings suggest a role for HSP27 in the balance between tumor dormancy and tumor progression, mediated by tumor-vascular interactions. Targeting HSP27 might offer a useful strategy in cancer treatment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Naumov GN, Folkman J, Straume O. Tumor dormancy due to failure of angiogenesis: Role of the microenvironment. Clin Exp Metastasis. 2009;26:51–60. - PubMed
-
- Black WC, Welch HG. Advances in diagnostic imaging and overestimations of disease prevalence and the benefits of therapy. N Engl J Med. 1993;328:1237–1243. - PubMed
-
- Folkman J, Kalluri R. Cancer without disease. Nature. 2004;427:787. - PubMed
-
- Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996;86:353–364. - PubMed
-
- Naumov GN, Akslen LA, Folkman J. Role of angiogenesis in human tumor dormancy: Animal models of the angiogenic switch. Cell Cycle. 2006;5:1779–1787. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
