

Epigenetic changes in pediatric solid tumors: promising new targets
- PMID: 22589485
- PMCID: PMC3691809
- DOI: 10.1158/1078-0432.CCR-11-1921
Epigenetic changes in pediatric solid tumors: promising new targets
Abstract
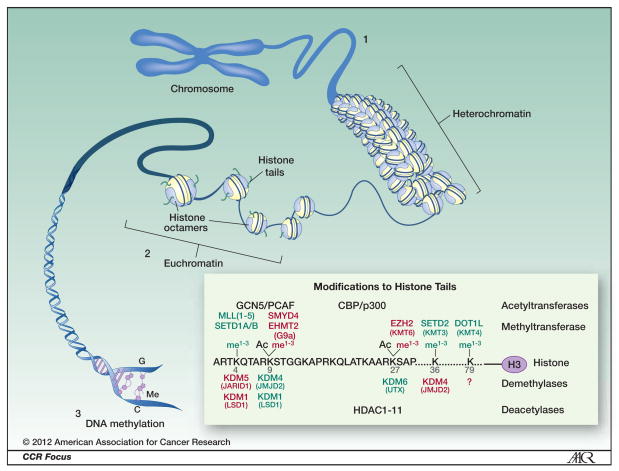
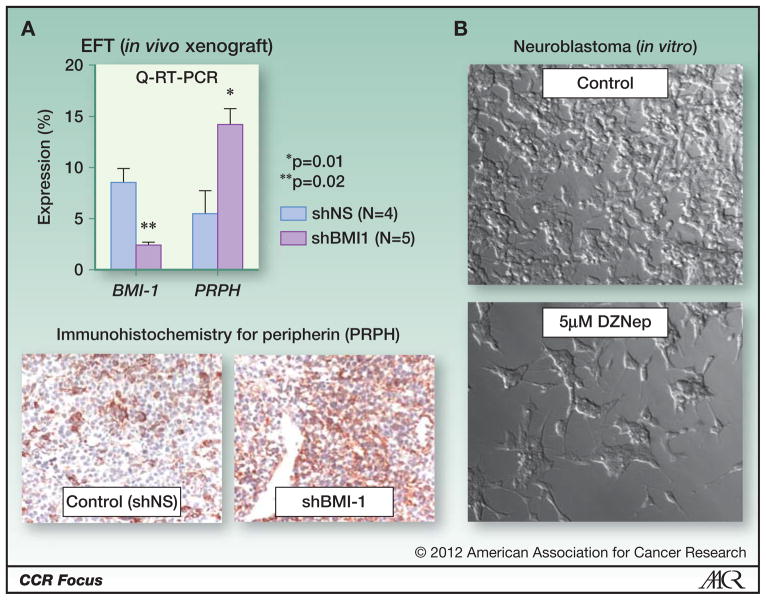
Cancer is being reinterpreted in the light of recent discoveries related to the histone code and the dynamic nature of epigenetic regulation and control of gene programs during development, as well as insights gained from whole cancer genome sequencing. Somatic mutations in or deregulated expression of genes that encode chromatin-modifying enzymes are being identified with high frequency. Nowhere is this more relevant than in pediatric embryonal solid tumors. A picture is emerging that shows that classic genetic alterations associated with these tumors ultimately converge on the epigenome to dysregulate developmental programs. In this review, we relate how alterations in components of the transcriptional machinery and chromatin modifier genes contribute to the initiation and progression of pediatric solid tumors. We also discuss how dramatic progress in our understanding of the fundamental mechanisms that contribute to epigenetic deregulation in cancer is providing novel avenues for targeted cancer therapy.
©2012 AACR.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
