

Entry of herpes simplex virus type 1 (HSV-1) into the distal axons of trigeminal neurons favors the onset of nonproductive, silent infection
- PMID: 22589716
- PMCID: PMC3349744
- DOI: 10.1371/journal.ppat.1002679
Entry of herpes simplex virus type 1 (HSV-1) into the distal axons of trigeminal neurons favors the onset of nonproductive, silent infection
Abstract
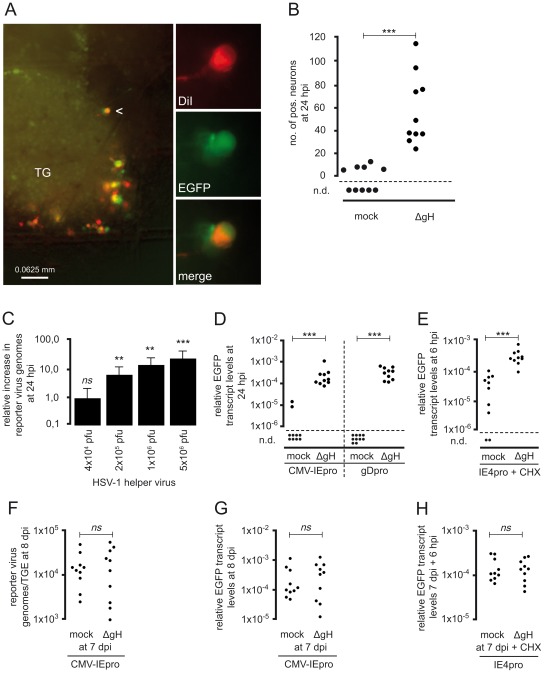
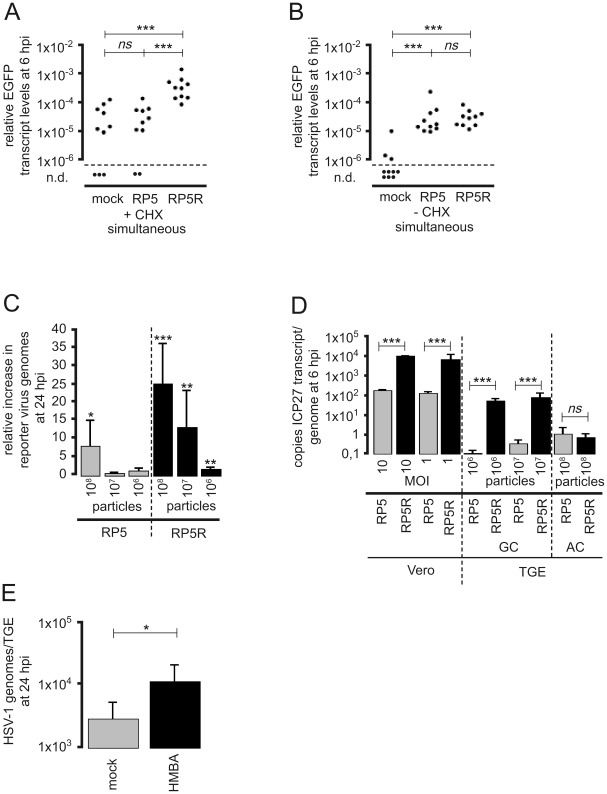
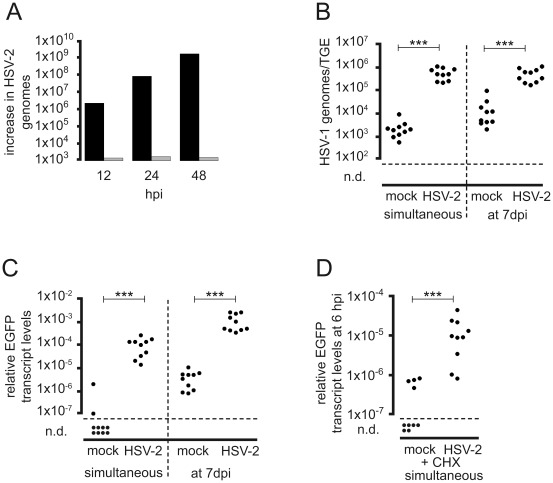
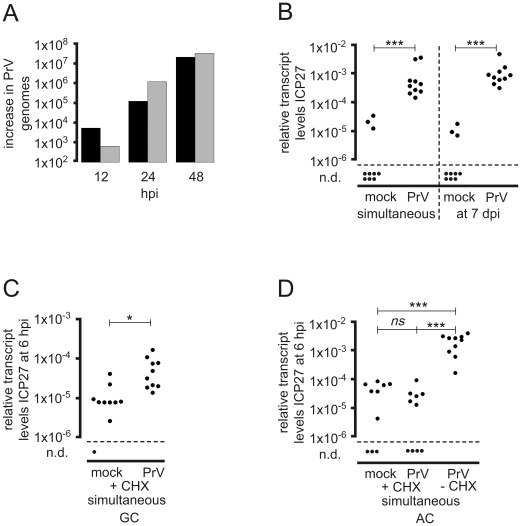
Following productive, lytic infection in epithelia, herpes simplex virus type 1 (HSV-1) establishes a lifelong latent infection in sensory neurons that is interrupted by episodes of reactivation. In order to better understand what triggers this lytic/latent decision in neurons, we set up an organotypic model based on chicken embryonic trigeminal ganglia explants (TGEs) in a double chamber system. Adding HSV-1 to the ganglion compartment (GC) resulted in a productive infection in the explants. By contrast, selective application of the virus to distal axons led to a largely nonproductive infection that was characterized by the poor expression of lytic genes and the presence of high levels of the 2.0-kb major latency-associated transcript (LAT) RNA. Treatment of the explants with the immediate-early (IE) gene transcriptional inducer hexamethylene bisacetamide, and simultaneous co-infection of the GC with HSV-1, herpes simplex virus type 2 (HSV-2) or pseudorabies virus (PrV) helper virus significantly enhanced the ability of HSV-1 to productively infect sensory neurons upon axonal entry. Helper-virus-induced transactivation of HSV-1 IE gene expression in axonally-infected TGEs in the absence of de novo protein synthesis was dependent on the presence of functional tegument protein VP16 in HSV-1 helper virus particles. After the establishment of a LAT-positive silent infection in TGEs, HSV-1 was refractory to transactivation by superinfection of the GC with HSV-1 but not with HSV-2 and PrV helper virus. In conclusion, the site of entry appears to be a critical determinant in the lytic/latent decision in sensory neurons. HSV-1 entry into distal axons results in an insufficient transactivation of IE gene expression and favors the establishment of a nonproductive, silent infection in trigeminal neurons.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
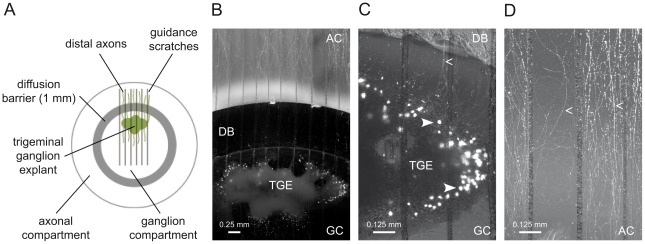
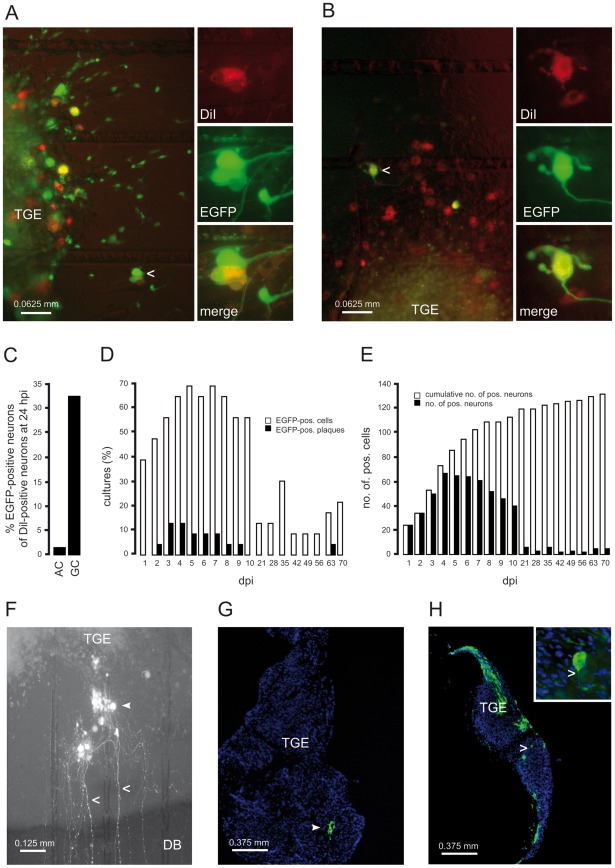
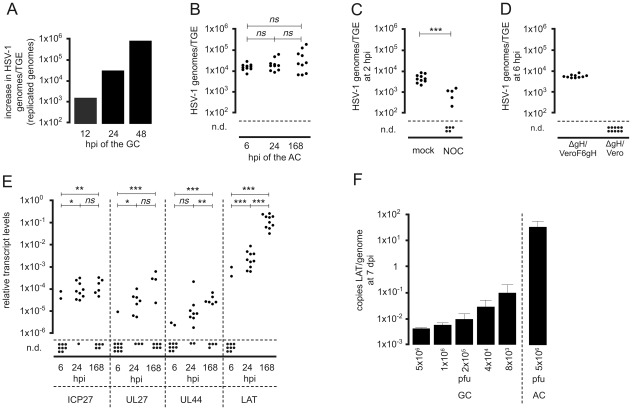
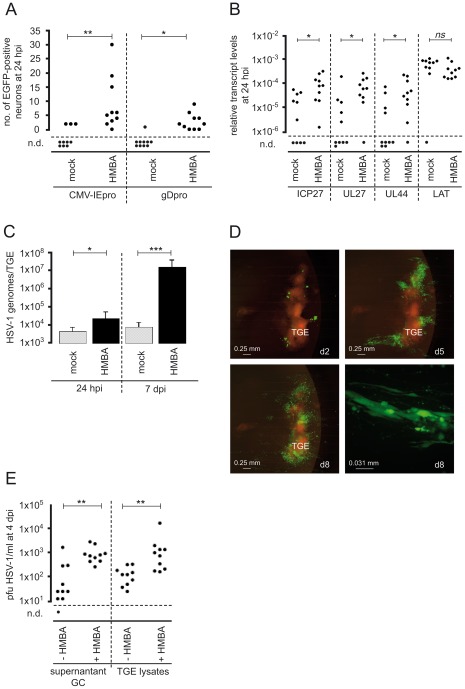
References
-
- Pellet PE, Roizman B. The familiy herpesviridae: A brief introduction. In: Fields BN, Knipe DM, Howley PM, editors. Fields Virology. 5 ed. New York, NY: Lippincott, Williams & Wilkins; 2007. pp. 2479–2500.
-
- Roizman B, Knipe DM, Whitley RJ. Herpes simplex viruses. In: Fields BN, Knipe DM, Howley PM, editors. Fields Virology. 5 ed. New York, NY: Lippincott, Williams & Wilkins; 2007. pp. 2501–2601.
-
- Diefenbach RJ, Miranda-Saksena M, Douglas MW, Cunningham AL. Transport and egress of herpes simplex virus in neurons. Rev Med Virol. 2008;18:35–51. - PubMed
-
- Preston CM, Efstathiou S. Molecular basis of HSV latency and reactivation. In: Arvin A, Campadelli-Fiume G, Mocarski E, Moore PS, Roizman B, et al., editors. Human Herpesviruses : Biology, Therapy, and Immunoprophylaxis. Cambridge: Cambridge University Press, PMID: 21348106; 2007. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
