

Maternal Wnt/β-catenin signaling coactivates transcription through NF-κB binding sites during Xenopus axis formation
- PMID: 22590521
- PMCID: PMC3348924
- DOI: 10.1371/journal.pone.0036136
Maternal Wnt/β-catenin signaling coactivates transcription through NF-κB binding sites during Xenopus axis formation
Abstract
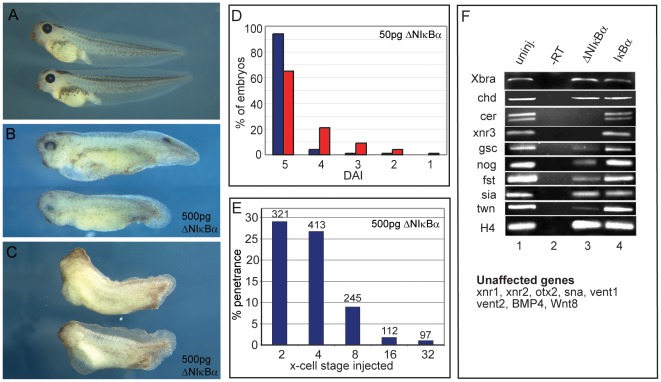
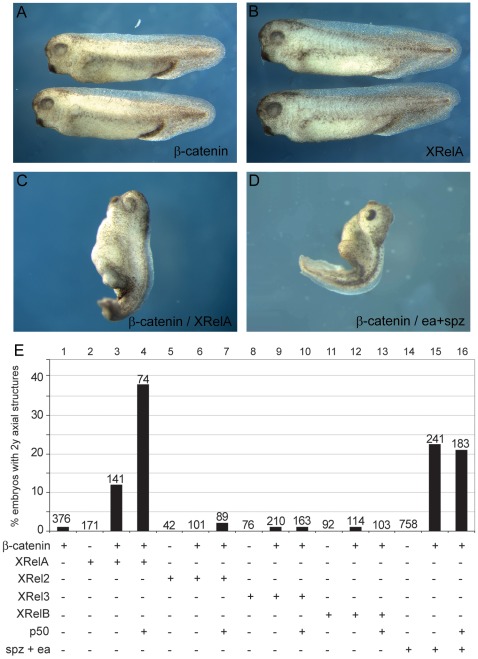
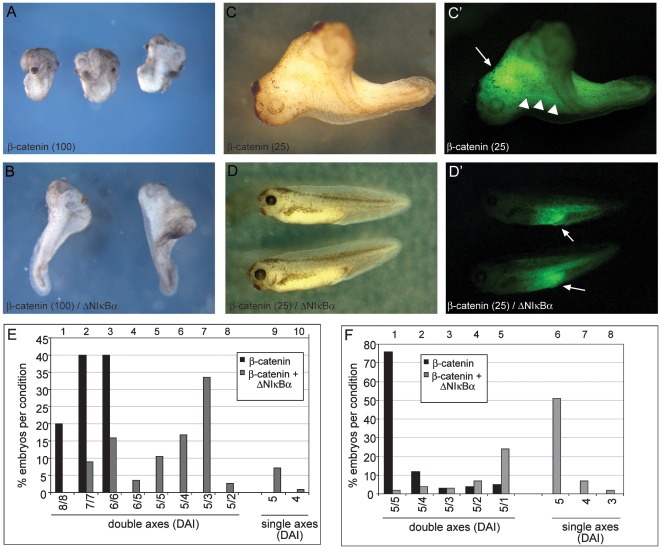
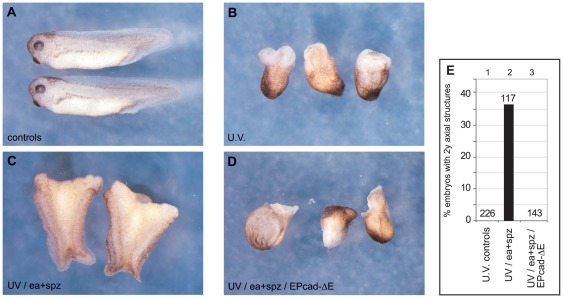
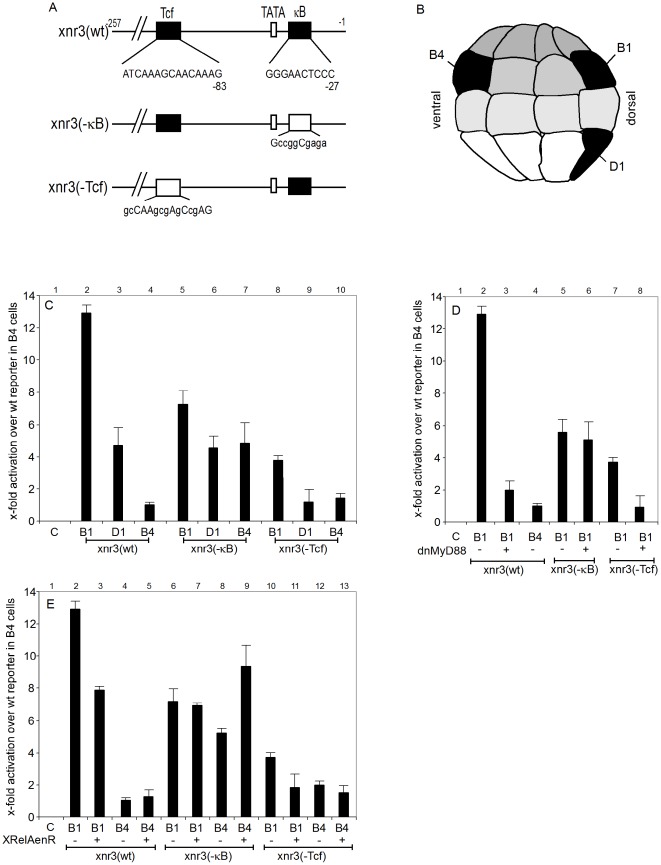
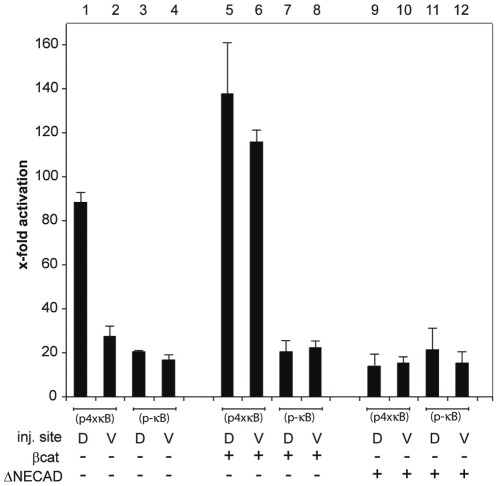
Maternal Wnt/β-Catenin signaling establishes a program of dorsal-specific gene expression required for axial patterning in Xenopus. We previously reported that a subset of dorsally expressed genes depends not only on Wnt/β-Catenin stimulation, but also on a MyD88-dependent Toll-like receptor/IL1-receptor (TLR/IL1-R) signaling pathway. Here we show that these two signal transduction cascades converge in the nucleus to coactivate gene transcription in blastulae through a direct interaction between β-Catenin and NF-κB proteins. A transdominant inhibitor of NF-κB, ΔNIκBα, phenocopies loss of MyD88 protein function, implicating Rel/NF-κB proteins as selective activators of dorsal-specific gene expression. Sensitive axis formation assays in the embryo demonstrate that dorsalization by Wnt/β-Catenin requires NF-κB protein activity, and vice versa. Xenopus nodal-related 3 (Xnr3) is one of the genes with dual β-Catenin/NF-κB input, and a proximal NF-κB consensus site contributes to the regional activity of its promoter. We demonstrate in vitro binding of Xenopus β-Catenin to several XRel proteins. This interaction is observed in vivo upon Wnt-stimulation. Finally, we show that a synthetic luciferase reporter gene responds to both endogenous and exogenous β-Catenin levels in an NF-κB motif dependent manner. These results suggest that β-Catenin acts as a transcriptional co-activator of NF-κB-dependent transcription in frog primary embryonic cells.
Conflict of interest statement
Figures

References
-
- Schneider S, Steinbeisser H, Warga RM, Hausen P. ß-Catenin translocation into nuclei demarcates the dorsalizing centers in frog and fish embryos. Mechanisms of Development. 1996;57:191–198. - PubMed
-
- Tao Q, Yokota C, Puck H, Kofron M, Birsoy B, et al. Maternal wnt11 activates the canonical wnt signaling pathway required for axis formation in Xenopus embryos. Cell. 2005;120:857–871. - PubMed
-
- Houston DW, Kofron M, Resnik E, Langland R, Destree O, et al. Repression of organizer genes in dorsal and ventral Xenopus cells mediated by maternal XTcf3. Development. 2002;129:4015–4025. - PubMed
-
- Standley HJ, Destree O, Kofron M, Wylie C, Heasman J. Maternal XTcf1 and XTcf4 have distinct roles in regulating Wnt target genes. Dev Biol. 2006;289:318–328. - PubMed
-
- Wardle FC, Smith JC. Transcriptional regulation of mesendoderm formation in Xenopus. Semin Cell Dev Biol. 2006;17:99–109. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
