

Identification of new agonists and antagonists of the insect odorant receptor co-receptor subunit
- PMID: 22590607
- PMCID: PMC3348135
- DOI: 10.1371/journal.pone.0036784
Identification of new agonists and antagonists of the insect odorant receptor co-receptor subunit
Abstract
Background: Insects detect attractive and aversive chemicals using several families of chemosensory receptors, including the OR family of olfactory receptors, making these receptors appealing targets for the control of insects. Insect ORs are odorant-gated ion channels, comprised of at least one common subunit (the odorant receptor co-receptor subunit, Orco) and at least one variable odorant specificity subunit. Each of the many ORs of an insect species is activated or inhibited by an unique set of odorants that interact with the variable odorant specificity subunits, making the development of OR directed insect control agents complex and laborious. However, several N-,2-substituted triazolothioacetamide compounds (VUAA1, VU0450667 and VU0183254) were recently shown to act directly on the highly conserved Orco subunit, suggesting that broadly active compounds can be developed. We have explored the chemical space around the VUAA1 structure in order to identify new Orco ligands.
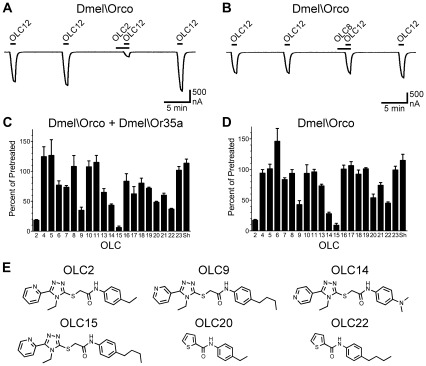
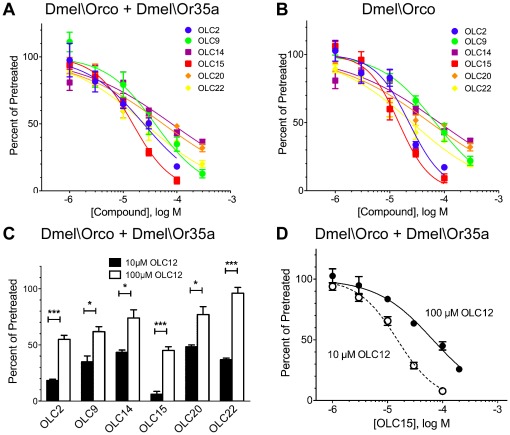
Principal findings: We screened ORs from several insect species, using heterologous expression in Xenopus oocytes and an electrophysiological assay, with a panel of 22 compounds structurally related to VUAA1. By varying the nitrogen position in the pyridine ring and altering the moieties decorating the phenyl ring, we identified two new agonists and a series of competitive antagonists. Screening smaller compounds, similar to portions of the VUAA1 structure, also yielded competitive antagonists. Importantly, we show that Orco antagonists inhibit odorant activation of ORs from several insect species. Detailed examination of one antagonist demonstrated inhibition to be through a non-competitive mechanism.
Conclusions: A similar pattern of agonist and antagonist sensitivity displayed by Orco subunits from different species suggests a highly conserved binding site structure. The susceptibility to inhibition of odorant activation by Orco antagonism is conserved across disparate insect species, suggesing the ligand binding site on Orco as a promising target for the development of novel, broadly active insect repellants.
Conflict of interest statement
Figures



References
-
- Hallem EA, Dahanukar A, Carlson JR. Insect odor and taste receptors. Annu Rev Entomol. 2006;51:113–135. - PubMed
-
- Vosshall LB, Stocker RF. Molecular architecture of smell and taste in Drosophila. Annu Rev Neurosci. 2007;30:505–533. - PubMed
-
- Sato K, Pellegrino M, Nakagawa T, Vosshall LB, Touhara K. Insect olfactory receptors are heteromeric ligand-gated ion channels. Nature. 2008;452:1002–1006. - PubMed
-
- Wicher D, Schafer R, Bauernfeind R, Stensmyr MC, Heller R, et al. Drosophila odorant receptors are both ligand-gated and cyclic-nucleotide-activated cation channels. Nature. 2008;452:1007–1011. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
