

Sister chromatid cohesion establishment occurs in concert with lagging strand synthesis
- PMID: 22592531
- PMCID: PMC3368863
- DOI: 10.4161/cc.20547
Sister chromatid cohesion establishment occurs in concert with lagging strand synthesis
Abstract
Cohesion establishment is central to sister chromatid tethering reactions and requires Ctf7/Eco1-dependent acetylation of the cohesin subunit Smc3. Ctf7/Eco1 is essential during S phase, and a number of replication proteins (RFC complexes, PCNA and the DNA helicase Chl1) all play individual roles in sister chromatid cohesion. While the mechanism of cohesion establishment is largely unknown, a popular model is that Ctf7/Eco1 acetylates cohesins encountered by and located in front of the fork. In turn, acetylation is posited both to allow fork passage past cohesin barriers and convert cohesins to a state competent to capture subsequent production of sister chromatids. Here, we report evidence that challenges this pre-replicative cohesion establishment model. Our genetic and biochemical studies link Ctf7/Eco1 to the Okazaki fragment flap endonuclease, Fen1. We further report genetic and biochemical interactions between Fen1 and the cohesion-associated DNA helicase, Chl1. These results raise a new model wherein cohesin deposition and establishment occur in concert with lagging strand-processing events and in the presence of both sister chromatids.
Figures
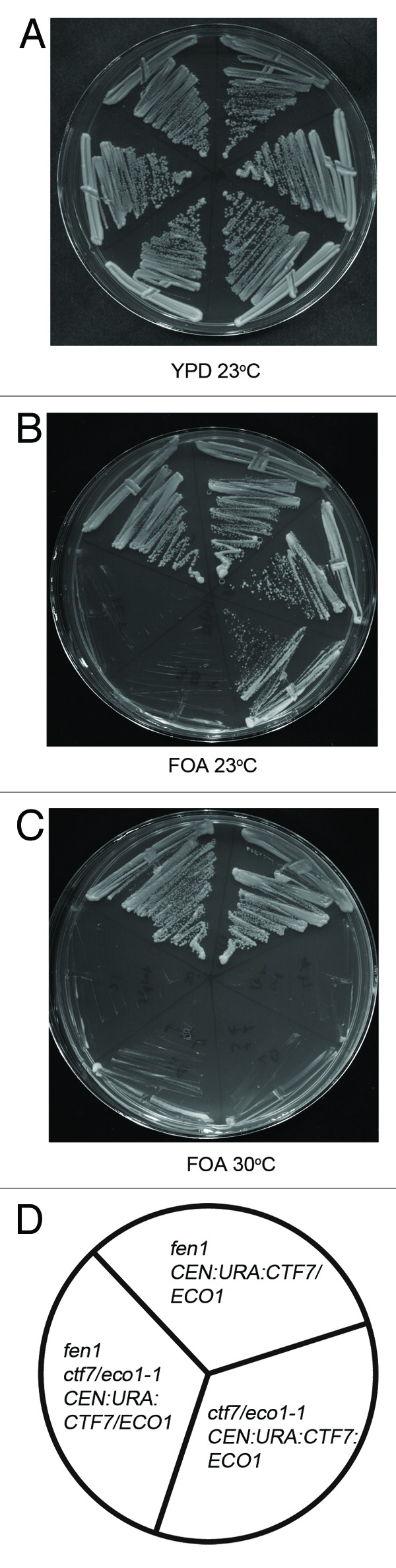
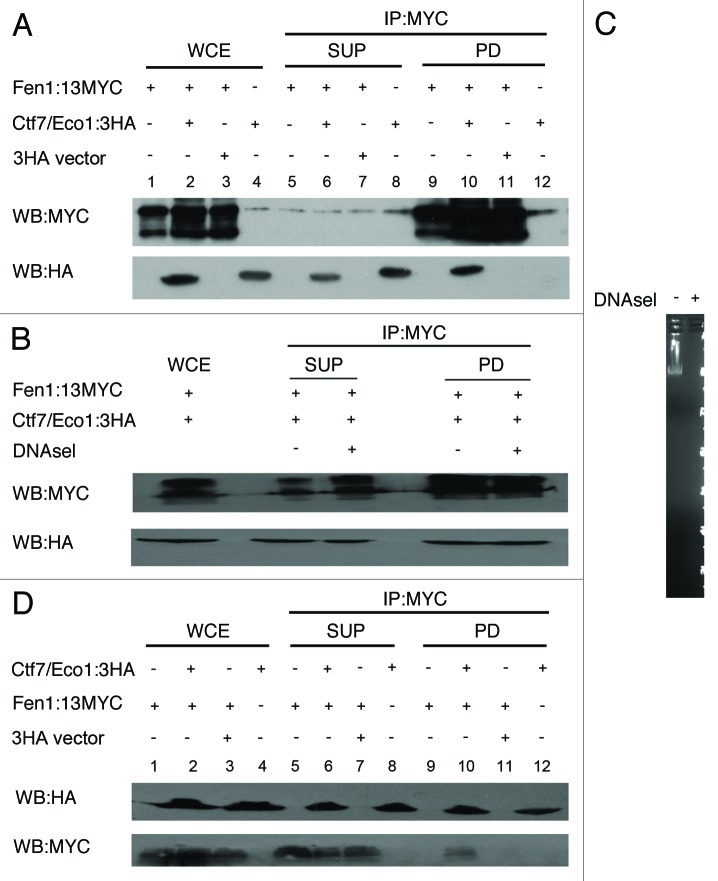
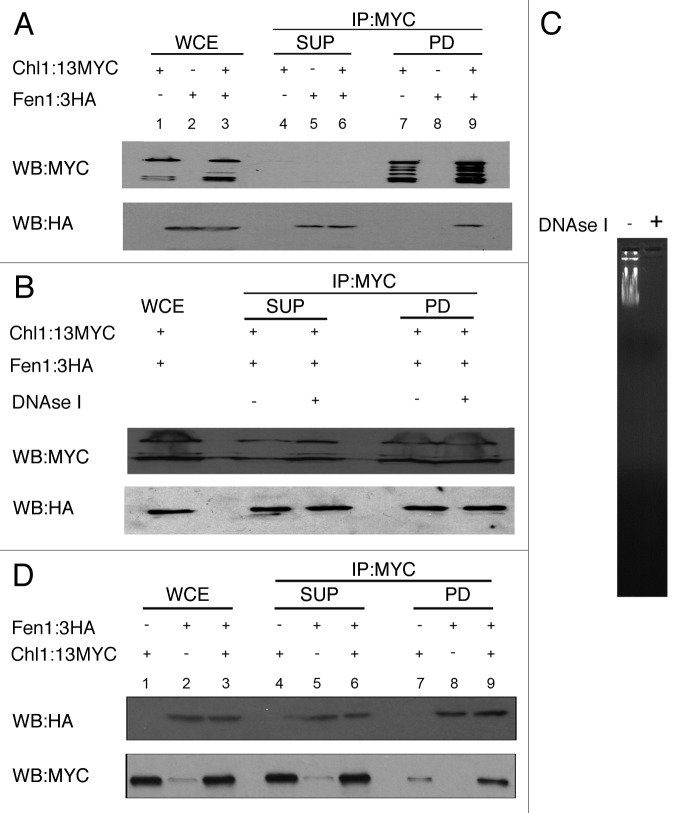
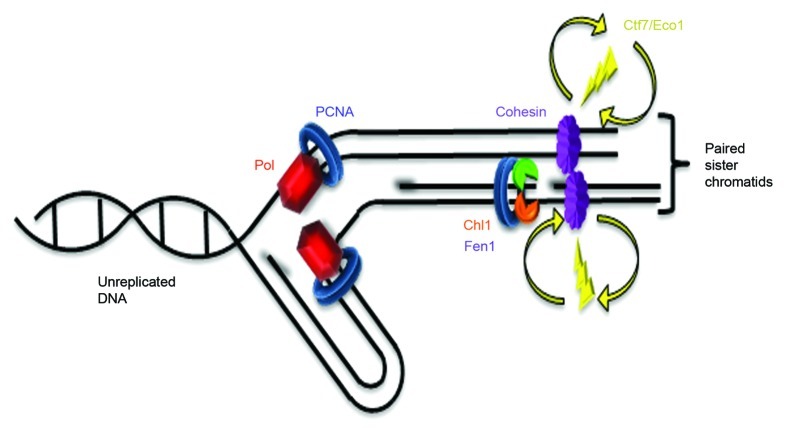
Comment in
-
Setting the stage for cohesion establishment by the replication fork.Cell Cycle. 2012 Jun 15;11(12):2228-9. doi: 10.4161/cc.20962. Epub 2012 Jun 15. Cell Cycle. 2012. PMID: 22677705 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous