

Vpu mediates depletion of interferon regulatory factor 3 during HIV infection by a lysosome-dependent mechanism
- PMID: 22593165
- PMCID: PMC3421752
- DOI: 10.1128/JVI.00423-12
Vpu mediates depletion of interferon regulatory factor 3 during HIV infection by a lysosome-dependent mechanism
Abstract
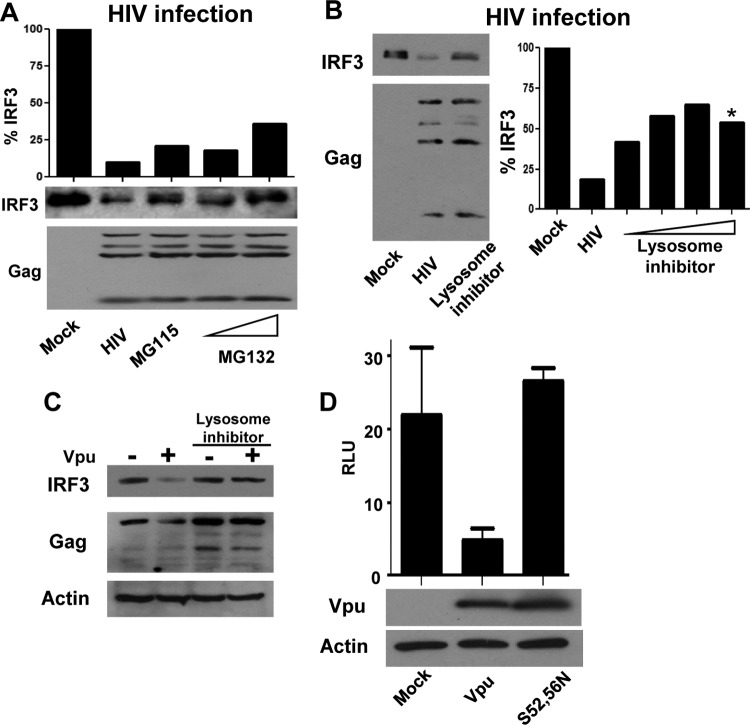
HIV has evolved sophisticated mechanisms to avoid restriction by intracellular innate immune defenses that otherwise serve to control acute viral infection and virus dissemination. Innate defenses are triggered when pattern recognition receptor (PRR) proteins of the host cell engage pathogen-associated molecule patterns (PAMPs) present in viral products. Interferon regulatory factor 3 (IRF3) plays a central role in PRR signaling of innate immunity to drive the expression of type I interferon (IFN) and interferon-stimulated genes (ISGs), including a variety of HIV restriction factors, that serve to limit viral replication directly and/or program adaptive immunity. Productive infection of T cells by HIV is dependent upon the targeted proteolysis of IRF3 that occurs through a virus-directed mechanism that results in suppression of innate immune defenses. However, the mechanisms by which HIV controls innate immune signaling and IRF3 function are not defined. Here, we examined the innate immune response induced by HIV strains identified through their differential control of PRR signaling. We identified viruses that, unlike typical circulating HIV strains, lack the ability to degrade IRF3. Our studies show that IRF3 regulation maps specifically to the HIV accessory protein Vpu. We define a molecular interaction between Vpu and IRF3 that redirects IRF3 to the endolysosome for proteolytic degradation, thus allowing HIV to avoid the innate antiviral immune response. Our studies reveal that Vpu is an important IRF3 regulator that supports acute HIV infection through innate immune suppression. These observations define the Vpu-IRF3 interface as a novel target for therapeutic strategies aimed at enhancing the immune response to HIV.
Figures
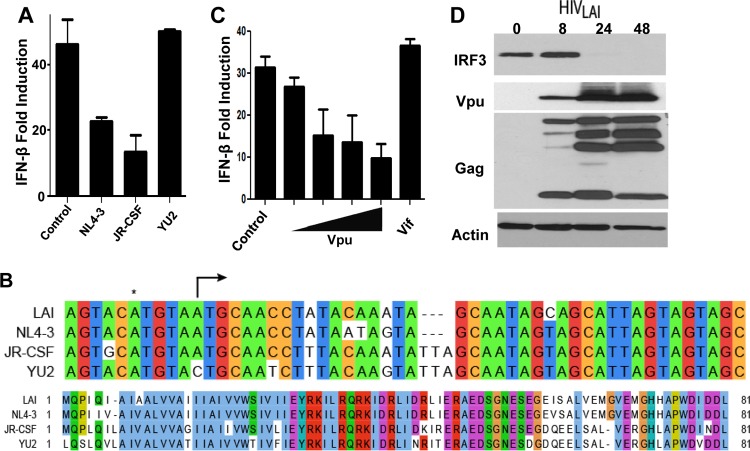
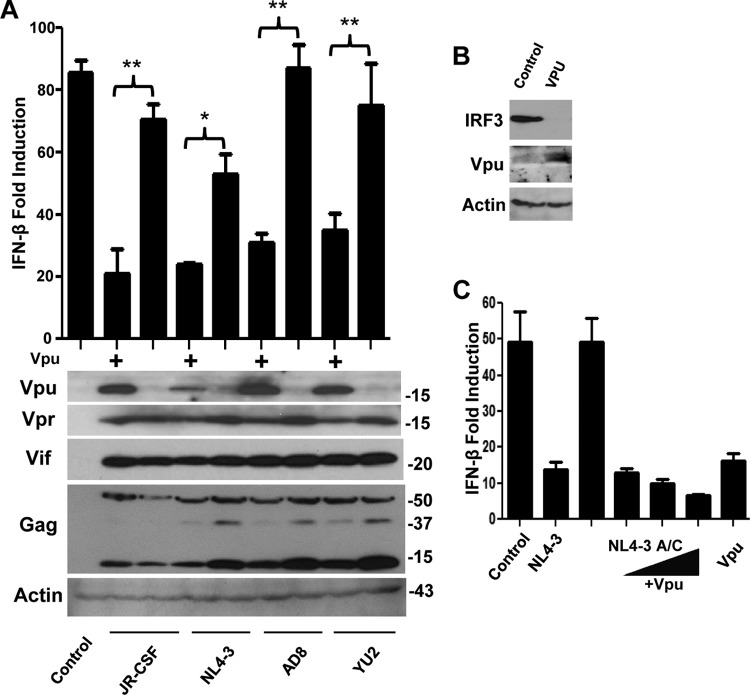
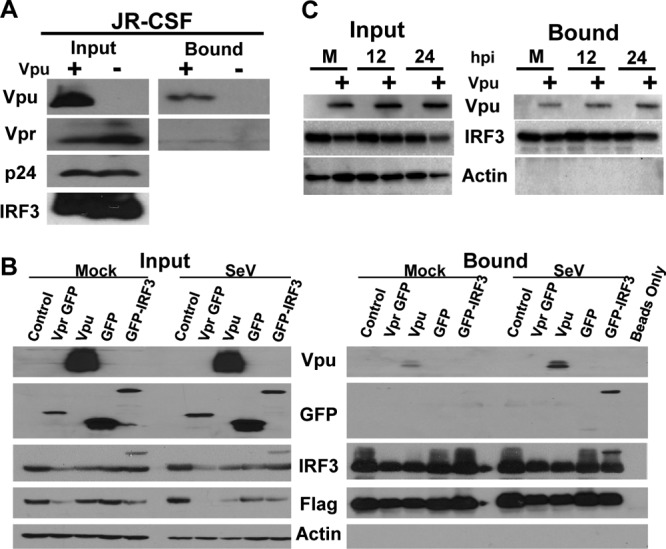
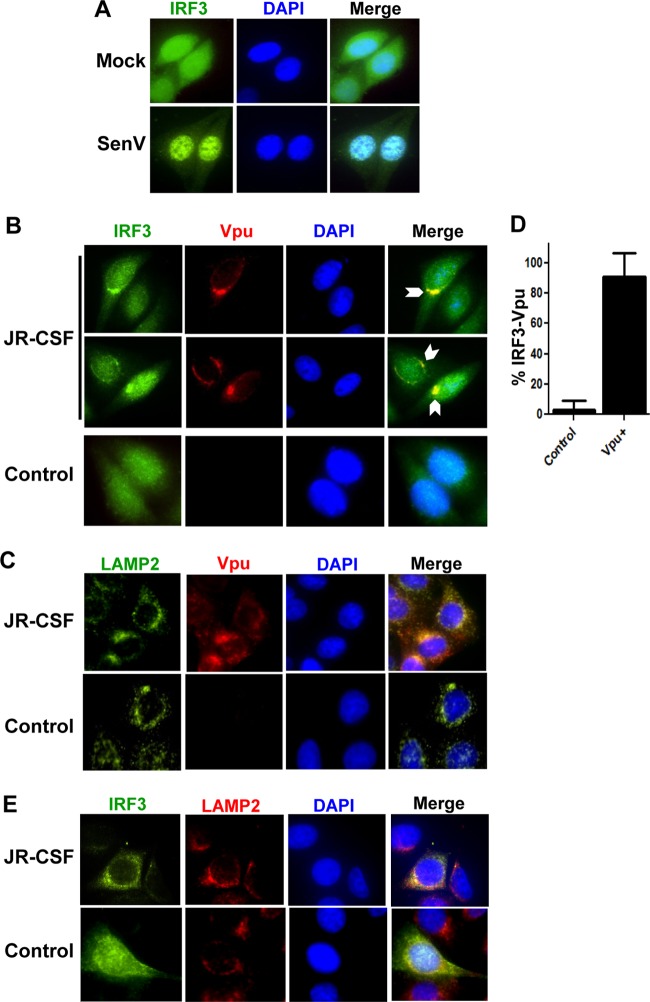
References
-
- Akira S, Uematsu S, Takeuchi O. 2006. Pathogen recognition and innate immunity. Cell 124:783–801 - PubMed
-
- Bosinger SE, et al. 2004. Gene expression profiling of host response in models of acute HIV infection. J. Immunol. 173:6858–6863 - PubMed
-
- Dejucq N, Simmons G, Clapham PR. 2000. T-cell line adaptation of human immunodeficiency virus type 1 strain SF162: effects on envelope, Vpu and macrophage-tropism. J. Gen. Virol. 81:2899–2904 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
